La inmunidad humoral es el principal mecanismo de defensa contra los microorganismos extracelulares y sus toxinas, en el cual, los componentes del sistema inmunitario que atacan a los antígenos, no son las células directamente sino los anticuerpos1 secretados por activación antigénica.
La respuesta efectora humoral está a cargo de las inmunoglobulinas o anticuerpos que son moléculas secretadas por células plasmáticas. Existen cinco clases de inmunoglobulinas: IgM, IgG, IgA, IgE e IgD, formadas por una unidad básica compuesta de dos cadenas polipetídicas globulares pesadas y dos cadenas livianas unidas entre sí por puentes disulfuro (A). Ambas cadenas presentan una zona constante (c) y una zona variable (v). En esta última, se encuentra una zona hipervariable formada por 10 a 15 aminoácidos que conforman el receptor idiotípico (r) responsable de la unión con el epitopo presente en el antígeno. Al ser sometida a digestión por papaína (flecha), esta molécula genera dos fragmentos, el fragmento Fab responsable de la unión con antígeno y el fragmento Fc que determina diversas funciones biológicas en las diferentes inmunoglobulinas. El fragmento Fab está formado por una región constante y una región variable de una cadena pesada y de una cadena liviana. El fragmento Fc está formado sólo por regiones constantes de cadenas pesadas. Las clases de inmunoglobulinas están determinadas por los diferentes isotipos de las cadenas pesadas. Estas pueden ser mu, gamma, alfa, delta o epsilon. Las cadenas livianas pueden ser kappa o bien lambda. Las distintas clases de inmunoglobulinas presentan diversas funciones biológicas.
os Anticuerpos desempeñan un papel importante en el sistema inmune. Las inmunoglobulinas presentes en la superficie del B-Linfocito envían en señales a los electores citoplásmicos y nucleares. Éstos también entregan el antígeno a la célula donde puede ser destruido, ser tramitado y ser vuelto a la superficie de la célula que se presentará por las moléculas de la clase II de MHC a las células de ayudante del fi c T del antígeno-speci.
Los linfocitos de T a su vez envían señales a las células de B para que maduren y reconozcan los antígenos y creen los anticuerpos apuntados específicamente contra él.
Inmunorespuesta Humoral Los Anticuerpos secretados por los linfocitos de B son responsables de la inmunorespuesta humoral. El sistema inmune humoral ayuda en la destrucción de patógeno externos y previene la extensión de infecciones intracelulares. Este sistema inmune también protege contra las toxinas.
Diversas funciones de diversas partes del anticuerpo Las dos porciones estructurales del anticuerpo, del constante (Fc) los fragmentos es decir variables (Fabuloso) y, comunican funciones biológicas distintas.
michelle m cordero rodriguez 2011-0288 Slide #1 La antigenicidad es la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. El lugar o determinante antigénico es la parte limitada de la molécula de un antígeno, adoptada especialmente a ciertas partes del anticuerpo específico: condiciona la aptitud del antígeno a combinarse selectivamente con el anticuerpo correspondiente. Un antígeno tiene generalmente varias localizaciones pudiendo unirse cada una de ellas con un anticuerpo específico diferente. Concepto relacionado: receptor de reconocimiento o de membrana, sensibilización y fragmento Fab.
Slide #2 Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos, receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los epítopos de las células T se presentan en la superficie de una célula presentadora de antígeno, concretamente a través de moléculas del MHC. Los epítopos de las células T presentados por moléculas MHC clase I son típicamente péptidos de entre 8 y 15 aminoácidos de longitud, mientras que las moléculas MHC clase II presentan péptidos más largos y las moléculas MHC no clásicas también presentan epítopos no peptídicos como los glucolípidos. Los epítopos pueden ser trazados usando microarrays de proteínas, y con las técnicas ELISPOT o ELISA. Los epítopos tienen a veces reactividad cruzada. Esta propiedad es explotada por el sistema inmunitario en la regulación por anticuerpos anti-idiotípicos. Si un anticuerpo se une a un epítopo de un antígeno, el paratopo puede convertir el epítopo para otro anticuerpo que entonces se unirá a él. Si este segundo anticuerpo es de clase IgM, su unión puede acentuar la respuesta inmune; si el segundo anticuerpo es de clase IgG, su unión puede disminuirla. Actualmente se están realizando investigaciones intensivas para diseñar herramientas fiables que predecirán epítopos en proteínas.
michelle M. CORDERO rodriguez 2011-0288 Slide #3 Estructura de anticuerpo Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ. El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos. Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad. Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina. La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
Cadena ligera En los mamíferos hay dos tipos de cadena ligera, llamados lambda (λ) y kappa (κ). Una cadena ligera contiene dos dominios sucesivos: un dominio constante y un dominio variable. La longitud aproximada de la cadena ligera es de 211 a 217 aminoácidos. Cada anticuerpo contiene dos cadenas ligeras que son siempre idénticas. Sólo un tipo de cadena ligera, κ o λ, está presente dentro del mismo anticuerpo en mamíferos. Otros tipos de cadenas ligeras como la cadena iota (ι), se encuentran en los vertebrados inferiores como los condrictios y teleósteos.
Regiones Fab y Fc Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Slide #4 Tipos de anticuerpos Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla. El isotipo cambia durante el desarrollo y la activación de los linfocitos B. Antes de la maduración de estos últimos, cuando aún no se han expuesto a su antígeno, se conocen como linfocitos B vírgenes y sólo expresan el isotipo IgM en su forma anclada a la superficie celular. Los linfocitos comienzan a expresar tanto IgM como IgD cuando alcanzan la madurez y en ese momento están listos para responder a su antígeno. La activación de los linfocitos B sigue al encuentro y unión de éste con su antígeno, lo que estimula a la célula para que se divide y se diferencie en una célula productora de anticuerpos denominada plasmática. En esta forma activada, los linfocitos B comienzan a secretar anticuerpos en lugar de anclarlos a la membrana. Algunas células hijas de los linfocitos B activados sufren un cambio isotípico, un mecanismo que provoca que la producción de anticuerpos en las formas IgM o IgD se trasmute a los otros tipos, IgE, IgA o IgG, que desempeñan distintos papeles en el sistema inmunitario.
Slide #5 Inmunoglobulinas valores normales
IgG : Elevadas en Mieloma por IgG, Sarcoidosis, Enfermedad Hepática crónica, enf. autoinmunes, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgG. Su valor normal es 700-1500 mg/dl.
IgM : Elevadas en : Mieloma por IgM, Enfermedad Hepática crónica, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgM. Su nivel normal es 60-300 mg/dl.
IgA: Elevada en Mieloma por IgA, Cirrosis Hepática, infecciones crónicas, artritis reumatoidea, LES. Disminuida en Inmunodeficiencias adquiridas, Mieloma no por IgA. Su valor normal es 60-400 mg/dl.
IgE: Elevada en enfermedades atópicas, asma, fiebre del heno, infecciones parasitarias, Mieloma por IgE. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgE. Su valor norma es 0.002-0.2 mg/dl
IgD: Elevada en Infecciones crónicas, Mieloma por IgD, enfermedades autoinmunes. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgD. Su valor normal es 0-14 mg/dl
Slide #6 michelle m cordero 2011-0288 La inmunoglobulina G (IgG) es una de las cinco clases de anticuerpos humorales producidos por el organismo. Se trata de la inmunoglobulina predominante en los fluidos internos del cuerpo, como son la sangre, el líquido cefalorraquídeo y el liquido peritoneal (líquido presente en la cavidad abdominal). Esta proteína especializada es sintetizada por el organismo en respuesta a la invasión de bacterias, hongos y virus. Es la inmunoglobulina más abundante del suero, con una concentración de 600-1.800 mg por 100 mL.1 La IgG constituye el 80% de las inmunoglobulinas totales. La IgG es la única clase de inmunoglobulinas que atraviesa la placenta, transmitiendo la inmunidad de la madre al feto. Es la inmunoglobulina más pequeña, con un peso molecular de 150.000 Daltons, así puede pasar fácilmente del sistema circulatorio del cuerpo a los tejidos. La síntesis de IgG se controla principalmente por el estímulo de los antígenos. En el caso de animales axénicos (sin microbios), con niveles de IgG muy bajos, el nivel de IgG se eleva en cuanto se les traslada a un ambiente normal. Se sabe que no todas las Inmunoglobulinas de una misma clase tienen idéntica estructura, sin que dentro de las clases se pueden establecer subclases considerando la secuencia de aminoácidos de la región constante de las cadenas H y el diferente número y situación de los puentes disulfuro intercatenarios establecidos entre las cadenas pesadas. Así, la IgG humana se divide en cuatro subclases (IgG1, IgG2, IgG3 e IgG4) y la IgA e IgM en dos (IgA1 e IgA2; IgM1 e IgM2) respectivamente.
Slide #7 IgM La inmunoglobulina M (IgM) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo, constituyendo un 6% de la población presente en sangre. Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso,macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons), aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM. La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizardeterminados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
Slide #8 michelle m cordero 2011-0288 La inmunoglobulina A (IgA) es la clase predominante de anticuerpo en las secreciones sero mucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica. (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 mL.
Slide #9 Las respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
Slide #10 La inmunoglobulina D (IgD) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo humano. Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma. Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina. La IgD es una proteína funcional importante, pero su papel no es bien conocido; se sugiere que es un receptor antigénico de membrana, que conduciría a la diferenciación linfocitaria y también que es un ligando para los receptores de IgD en la inmunoregulación de las células T ‘helper’. Es muy susceptible a la proteólisis. La IgD representa menos del 1% del total de inmunoglobulinas plasmáticas; la concentración en suero depende de la edad y de la herencia genética. En suero de pacientes con mieloma IgD se encuentran concentraciones de IgD muy elevadas.
Slide #11 michelle m cordero 2011-0288 La IgE es una glicoproteína de aproximadamente 190 kDa. Como otros anticuerpos monoméricos, está compuesta de dos cadenas pesadas y dos ligeras, unidas por puentes disulfuro. Su cadena pesada consta de 5 dominios, uno variable y cuatro constantes de tipo ε. La mayor parte de la IgE se encuentra unida a FcεRI en la superficie de los mastocitos, eosinófilos y basófilos. El reconocimiento de un antígeno por la IgE desencadena complejas reacciones inmunitarias, entre las que pueden destacarse, por ejemplo, la desgranulación de los mastocitos, que liberan sustancias vasoactivas como la histamina, así como la intervención de los eosinófilos en la respuesta inflamatoria. Contiene de un 10 a un 12% en peso de glúcidos y su concentración en el suero es de 0,01 a 0,10 mg por 100 mL. La producción de IgE específica de antígenos no patogénicos, como los encontrados en fármacos, alimentos, y alérgenos estacionales, puede provocar reacciones alérgicas. La liberación del contenido de los gránulos de los mastocitos al reconocimiento de antígeno causa síntomas como la hinchazón de las vías aéreas, la rinitis, y la dermatitis. La IgE se fija a la superficie de membrana de las células cebadas o basofilas hasta por 2 semanas en una respuesta alérgica.
Slide 12 El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. La síntesis de las cadenas ligeras y pesadas se regula por genes que se encuentran en cromosomas distintos. Esto fue puesto de manifiesto empleando técnicas de digestión enzimática del DNA de células B y posterior hibridación con sondas de DNA complementario (cDNA), mediante la técnica de Southern Blot . Mediante esta técnica, se pudo observar que usando un cDNA marcado radioactivamente como sonda correspondiente a parte de una cadena pesada, éste se unía a segmentos de DNA de líneas celulares de ratón que contenían el cromosoma 12. Por otra parte, empleando igualmente sondas de cDNA marcadas radioactivamente frente al DNA de cadenas ligeras de tipo k, éstas se unen a células que contienen el cromosoma 6, mientras que las sondas para DNA de cadenas ligeras l lo hacen alcromosoma 16. Cuando una persona es alérgica a una sustancia en particular, el sistema inmunitario cree, erróneamente, que está bajo una invasión antígenica por parásitos, y produce la IgE, en un intento de "proteger" el organismo; de esta manera, se inicia una cadena de acontecimientos que provocan los síntomas de la alergia. Si una persona sufre de asma producida por reacciones alérgicas, esta cadena de acontecimientos también derivará en síntomas de asma. La molécula de IgD tiene una larga región "bisagra" entre Fab y Fc, lo que proporciona a la molécula más susceptibilidad a la degradación proteolítica, así como una mayor flexibilidad que favorece la unión al antígeno, y presenta 3 dominios en la región constante.
Slide #13 Otra característica importante de la formación de inmunoglobulinas es que, en la síntesis de cada una de las cadenas de inmunoglobulinas participan varios segmentos de genes de cuyas combinaciones resultan los diversos genes funcionales, responsables directos de la codificación de cada una de las cadenas de inmunoglobulinas. Esto explica las múltiples posibilidades de formación del gran número de inmunoglobulinas partiendo de un limitado material genético. Estos segmentos codifican por separado la parte variable, la parte constante y las partes por las que ambas regiones se unen, esto es, las regiones bisagra. Los segmentos que codifican la parte variable son de dos tipos, segmentos V y segmentos D, responsables de la variabilidad y diversidad de las inmunoglobulinas respectivamente. Le siguen los segmentos de unión, responsables de unir los fragmentos anteriores con los segmentos C que codifican para la parte constante. El número de segmentos responsables de la codificación de la parte constante es muy limitado en relación con el número de segmentos que codifican la parte variable de las inmunoglobulinas.
Slide 14 En la síntesis de cadenas ligeras participan los segmentos V, J y C (es decir, no hay genes D para la cadena ligera). Así pues, en el proceso de recombinación de genes de cadenas ligeras se acopla un segmento V con un segmento J y el conjunto V/J se recombina con el segmento correspondiente a la parte constante C. El proceso de transcripción se hace de tal manera que el RNA mensajero contiene información secuenciada V, J y C. A su vez, un proceso similar, si bien esta vez en dos etapas, ocurre para la formación de una cadena pesada. En este caso participan los segmentos V, D, J, y C. Primero se produce la recombinación entre un segmento D y un segmento J. En la segunda fase, este conjunto D/J se recombina con un segmento V. El complejo V/D/J se puede por último recombinar con cada uno de los segmentos que codifican las regiones constantes, según la inmunoglobulina a formar. El proceso de recombinación con los genes C tiene lugar a nivel de RNA. Vamos a encontrar en este momento el siguiente orden: el segmento líder; un fragmento V/D/J reordenado y unido y a continuación cada uno de los genes que codifican para las regiones constantes (Cm; Cd; Ct, etc) separados entre sí por los correspondientes intrones y a su vez precedidos por sus respectivos promotores. Se piensa que el gen Cm es el situado en primer lugar en la secuencia de genes C, seguido de Cd. En células B en reposo, el único y largo trascrito primario de RNA va a contener la información del complejo V/D/J más la correspondiente a los genes Cm y Cd. Mediante mecanismos de procesamiento del RNA que analizaremos posteriormente, se va a proceder a la escisión en el trascrito primario de la información correspondiente a Cm o bien a Cd. Se da lugar de esta manera a una inmunoglobulina de tipo IgM o IgD, y este mecanismo es además el responsable de que en las células B en reposo se encuentre la expresión simultánea de ambas inmunoglobulinas.
Slide #15 michelle m cordero 2011-0288 El calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. El calostro de todas las especies mamíferas contiene ingredientes inmunizantes contra las bacterias Escherichia coli, que pueden llegar a causar diarrea y vómito. Además también neutralizan toxinas, virus y las bacterias en el sistema linfático y circulatorio. Se ha llegado a comprobar, que en las 72 h posteriores al alumbramiento, la vaca elimina hasta 2 kg de inmunoglobulinas.
michelle m. cordero 2011-0288 Slide# 16 Funciones de los Anticuerpos Puesto que los anticuerpos se dan de forma libre en el torrente sanguíneo, se dice que son parte del sistema inmunitario humoral. Los anticuerpos circulantes son producidos por líneas clonales de linfocitos B que responden específicamente a un antígeno que puede ser un fragmento de proteína de la cápside viral, por ejemplo. Los anticuerpos contribuyen a la inmunidad de tres formas distintas: pueden impedir que los patógenos entren en las células o las dañen al unirse a ellas (neutralización). Pueden estimular la eliminación de un patógeno por losmacrófagos y otras células revistiendo al patógeno (opsonización) y pueden desencadenar la destrucción directa del patógeno estimulando otrasrespuestas inmunes como la vía del complemento (lisis). Activación del complemento Los anticuerpos que se unen a la superficie de los antígenos, por ejemplo, en una bacteria, atraen los primeros componentes de la cascada del complemento mediante su región Fc e inician la activación del sistema "clásico" del complemento. Esto acaba con la muerte de la bacteria de dos formas: Primero, la unión de las moléculas del complemento con el anticuerpo marca al microbio para la ingestión por los fagocitos en un proceso llamado opsonización. Estos fagocitos son atraídos por ciertas moléculas del complemento. En segundo lugar, algunos componentes del sistema del complemento forman un complejo de ataque a membrana para ayudar a los anticuerpos a matar a la bacteria por medio de lisis. Los anticuerpos más efectivos en la activación del Sistema del Complemento son los de tipo IgM y los de tipo IgG subclase 1 y 3 (IgG1 e IgG3).
michelle m cordero 2011-0288 Activación de células efectoras Para combatir a los patógenos que se replican en el exterior de las células, los anticuerpos se unen a los patógenos para ensamblarlos juntos provocando su aglutinación. Puesto que un anticuerpo tiene al menos dos paratopos se puede unir a más de un antígeno acoplándose a epítopos idénticos portados en las superficies de esos antígenos. Revistiendo al patógeno, los anticuerpos estimulan las funciones efectoras contra éste en las células que reconocen la región Fc. Aquellas células que reconocen los patógenos revestidos tienen receptores del Fc que, como su nombre indica, interactúan con la región Fc de los anticuerpos IgA, IgG, e IgE. El acoplamiento de un anticuerpo particular con el receptor Fc de una determinada célula desencadena en ella una función efectora: los fagocitos realizarán lafagocitosis, las células cebadas y los neutrófilos producirán la degranulación, las células asesinas naturales liberarán citoquinas y moléculas citotóxicas que finalmente acabarán con la destrucción del microbio invasor. Los receptores Fc son específicos del isotipo, lo que da una mayor flexibilidad al sistema inmune, afectando solo al mecanismo inmune adecuado para los distintos patógenos.
Slide# 17 Activación células B La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T. Activación dependiente de células T Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie. La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores
michelle m cordero 2011-0288 para la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipo IgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes. Activación T-independiente Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes: • Tipo 1, o activación policlonal T-independiente; • Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.
jose morales 86555 La Inmunidad Humoral es aquella que se caracteriza porque los principales componentes del sistema Inmunológico, los Anticuerpos o Inmunoglobulinas bloquean o lisan Noxas biológicas en forma directa en el medio extracelular, es decir, sin haber ingresado a las células eucariotas del propio cuerpo, no hay participación directa de células Inmunológicas(Leucocitos-Linfocitos) sino solo los Anticuerpos sintetizados por los Linfocitos B y T.
En cambio la Inmunidad Celular es aquella en la cual participan directamente los Linfocitos T, bloqueando la proliferación de las Noxas Biológicas en el interior de las propias células eucariotas del cuerpo infectadas por Virus y Bacterias a la cual no tienen ecceso directo los Anticuerpos.
Se destacan los Linfocitos T Citotóxicos o Citolíticos que vuelcan en la membrana plasmática de las células del cuerpo infectadas Linfocinas, Citoquininas produciendo la Lisis de la propia célula infectada dejando en libertad a las Noxas para ser fagocitadas por otras células inmunológicas(Neutrófilos-Monocitos), las células T Cooperadoras que ayudan a los Linfocitos B y T Citotóxicos y las células Supresoras que se especializan en la activación o inhibición de las células B y T, aumentando su proliferación, Inhibiendo su proliferación o ambos procesos simultáneamente.
ALEJANDRA FIGUEROA 88099 Capacidad para dar lugar a la producción de anticuerpos. El grado de antigenicidad depende del tipo y cuantía de la sustancia concreta, de las circunstancias del huésped y de su grado de sensibilidad al antígeno y de su capacidad para producir anticuerpos. Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos,1 receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los anticuerpos son proteínas plasmáticas globulares pesadas (~150 kDa), también conocidas como inmunoglobulinas. Tienen cadenas de azúcares unidas a alguno de sus residuos aminoácido.34 En otras palabras, los anticuerpos son glicoproteínas. La unidad básica funcional de cada anticuerpo es el monómero de inmunoglobulina, que contiene una sola unidad de Ig. Los anticuerpos secretados también pueden ser diméricos con dos unidades Ig, como en el caso de las IgA, tetraméricos con cuatro unidades Ig como en el caso de las IgM de teleósteo, o pentaméricos con cinco unidades de IgM, como en el caso de las IgM de mamíferos. El monómero de Ig es una molécula en forma de "Y" que consta de dos cadenas de polipéptido; dos cadenas pesadas idénticas y dos cadenas ligeras idénticas conectadas por enlaces disulfuro.31 Cada cadena se compone de dominios estructurales llamados dominios Ig. Estos dominios contienen entre 70 y 110 aminoácidos y se clasifican en diferentes categorías, por ejemplo en variables (IgV) y constantes (IgC) de acuerdo con su tamaño y función.38 Tienen un "pliegue inmunoglobulina" característico en el cual dos láminas beta generan una forma de "sándwich", permaneciendo juntas por interacciones entre cisteínas bien conservadas a lo largo de la evolución, así como otros aminoácidos cargados. Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ.3 El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos.3
1. Región Fab 2. Región Fc 3. Cadena pesada con un dominio variable (VH) seguido por un dominio constante (CH1), una región bisagra, y dos más constantes, los dominios (CH2 y CH3). 4. Cadena ligera con un dominio variable (VL) y uno constante (CL) 5. Lugar de unión al antígeno (paratopo) 6. Regiones bisagra. Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad.31 Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina.3 La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
ALEJANDRA FIGUEROA 88099 Los anticuerpos (también conocidos como inmunoglobulinas, abreviado Ig) son glicoproteínas del tipo gamma globulina. Pueden encontrarse de forma soluble en la sangre u otros fluidos corporales de los vertebrados, disponiendo de una forma idéntica que actúa como receptor de los linfocitos B y son empleados por el sistema inmunitario para identificar y neutralizar elementos extraños tales como bacterias, virus o parásitos.1 El anticuerpo típico está constituido por unidades estructurales básicas, cada una de ellas con dos grandes cadenas pesadas y dos cadenas ligeras de menor tamaño, que forman, por ejemplo, monómeros con una unidad, dímeros con dos unidades o pentámeros con cinco unidades. Los anticuerpos son sintetizados por un tipo de leucocito denominado linfocito B. Existen distintas modalidades de anticuerpo, isotipos, basadas en la forma de cadena pesada que posean. Se conocen cinco clases diferentes de isotipos en mamíferos que desempeñan funciones diferentes, contribuyendo a dirigir la respuesta inmune adecuada para cada distinto tipo de cuerpo extraño que encuentran Aunque la estructura general de todos los anticuerpos es muy semejante, una pequeña región del ápice de la proteína es extremadamente variable, lo cual permite la existencia de millones de anticuerpos, cada uno con un extremo ligeramente distinto. A esta parte de la proteína se la conoce como región hipervariable. Cada una de estas variantes se puede unir a una "diana" distinta, que es lo que se conoce como antígeno.3 Esta enorme diversidad de anticuerpos permite al sistema inmune reconocer una diversidad igualmente elevada de antígenos. La única parte del antígeno reconocida por el anticuerpo se denomina epítopo. Estos epítopos se unen con su anticuerpo en una interacción altamente específica que se denomina adaptación inducida, que permite a los anticuerpos identificar y unirse solamente a su antígeno único en medio de los millones de moléculas diferentes que componen un organismo. El reconocimiento de un antígeno por un anticuerpo lo marca para ser atacado por otras partes del sistema inmunitario. Los anticuerpos también pueden neutralizar sus objetivos directamente, mediante, por ejemplo, la unión a una porción de un patógeno necesaria para que éste provoque una infección.
ALEJANDRA FIGUEROA 88099 Los linfocitos B son los leucocitos de los cuales depende la inmunidad mediada por anticuerpos con actividad específica de fijación de antígenos. Las células B, que constituyen un 5 a 15% del total de linfocitos,[cita requerida] dan origen a las células plasmáticas que producen anticuerpos. Los linfocitos son de dos tipos principales, atendiendo a su origen y función: células T, que se diferencian inicialmente en el timo, y las células B, que se diferencian en el hígado y bazo fetal, y en la médula ósea del adulto (la 'B' proviene del latín Bursa fabricii, el órgano en el cual se desarrollan los linfocitos B en las aves). Durante su desarrollo, los linfocitos T y B adquieren receptores específicos para antígenos, el de las células B se le conoce como receptor de linfocito B (BCR). La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T. El calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. "Para el nacimiento del bebé se preparara uno de los alimentos más importantes de su vida, el calostro. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé.
Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ. El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos. Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad. Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina. La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
Cadena ligera En los mamíferos hay dos tipos de cadena ligera, llamados lambda (λ) y kappa (κ). Una cadena ligera contiene dos dominios sucesivos: un dominio constante y un dominio variable. La longitud aproximada de la cadena ligera es de 211 a 217 aminoácidos. Cada anticuerpo contiene dos cadenas ligeras que son siempre idénticas. Sólo un tipo de cadena ligera, κ o λ, está presente dentro del mismo anticuerpo en mamíferos. Otros tipos de cadenas ligeras como la cadena iota (ι), se encuentran en los vertebrados inferiores como los condrictios y teleósteos.
Regiones Fab y Fc Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Respuesta Inmune Humoral La ausencia de este tipo de respuesta deja al individuo tan indefenso frente a toda clase de gérmenes patógenos y otras agresiones, que es incompatible con la vida si no se instaura a tiempo un tratamiento adecuado. La respuesta inmune humoral es mediatizada por los linfocitos B, que como se ha dicho anteriormente reconocen al antigeno a través de las inmunoglobulinas de membrana. Sin embargo este estimulo no es suficiente para que se inicien los procesos de proliferación de estas células. Para ello es necesario que los linfocitos B además del estimulo antigénico reciban el estimulo de ciertas interleucinas. El elemento efector final de la respuesta humoral son las inmunoglobulinas. El termino inmunoglobulina fue propuesto por Heberman para designar a todas las sustancias con capacidad de anticuerpo, esto es con capacidad de anteponerse al antigeno. Las inmunoglobulinas son de cinco clases: inmunoglobulina M (IgM), inmunoglobulina A (IgA), inmunoglobulina G (IgG), inmunoglobulina D (IgD) e inmunoglobulina E (IgE). Las inmunoglobulinas tienen la propiedad de unirse específicamente al antígeno que indujo su formación. Tras la unión antígeno-anticuerpo (Ag-Ac), las sustancias extrañas (o antígenos) son destruídas por las inmunoglobulinas a través de mecanismos, que pueden ser diferentes según el tipo de inmunoglobulina que participa. Esto se debe a que aunque las distintas clases de inmunoglobulinas tienen una estrucutura igual en ciertas partes de la molecula, en otras partes presentan una estrucura distinta. Podemos decir que las inmunoglobulinas, al detectar al antígeno y unirse a el, actúan como transductores de la información de la presencia de los mismos, que serán posteriormente destruidos por el mecanismo más idoneo, en el que colaborarán además del propio anticuerpo el sistema del complemento, macrófagos, los polimorfonucleares o células K. El término complemento engloba, una gran variedad de proteinas, que interactúan en un determinado orden, se representan por C' y se encuentran en el suero. Cuando se produce la activación del C' se pone en marcha una serie de reacciones, en forma de "cascada", de tal forma que se van generando productos activos que además de influir en que la reacción prosiga tienen diferentes acciones biológicas importantes en la defensa del organismo.
88347 La antigenicidad es la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. El lugar o determinante antigénico es la parte limitada de la molécula de un antígeno, adoptada especialmente a ciertas partes del anticuerpo específico: condiciona la aptitud del antígeno a combinarse selectivamente con el anticuerpo correspondiente. Un antígeno tiene generalmente varias localizaciones pudiendo unirse cada una de ellas con un anticuerpo específico diferente. Concepto relacionado: receptor de reconocimiento o de membrana, sensibilización y fragmento Fab. Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos, receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los epítopos de las células T se presentan en la superficie de una célula presentadora de antígeno, concretamente a través de moléculas del MHC. Los epítopos de las células T presentados por moléculas MHC clase I son típicamente péptidos de entre 8 y 15 aminoácidos de longitud, mientras que las moléculas MHC clase II presentan péptidos más largos y las moléculas MHC no clásicas también presentan epítopos no peptídicos como los glucolípidos. Los epítopos pueden ser trazados usando microarrays de proteínas, y con las técnicas ELISPOT o ELISA. Los epítopos tienen a veces reactividad cruzada. Esta propiedad es explotada por el sistema inmunitario en la regulación por anticuerpos anti-idiotípicos. Si un anticuerpo se une a un epítopo de un antígeno, el paratopo puede convertir el epítopo para otro anticuerpo que entonces se unirá a él. Si este segundo anticuerpo es de clase IgM, su unión puede acentuar la respuesta inmune; si el segundo anticuerpo es de clase IgG, su unión puede disminuirla. Actualmente se están realizando investigaciones intensivas para diseñar herramientas fiables que predecirán epítopos en proteínas. Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ. El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos. Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad. Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina. La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
Cadena ligera En los mamíferos hay dos tipos de cadena ligera, llamados lambda (λ) y kappa (κ). Una cadena ligera contiene dos dominios sucesivos: un dominio constante y un dominio variable. La longitud aproximada de la cadena ligera es de 211 a 217 aminoácidos. Cada anticuerpo contiene dos cadenas ligeras que son siempre idénticas. Sólo un tipo de cadena ligera, κ o λ, está presente dentro del mismo anticuerpo en mamíferos. Otros tipos de cadenas ligeras como la cadena iota (ι), se encuentran en los vertebrados inferiores como los condrictios y teleósteos.
88347 Regiones Fab y Fc Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla. El isotipo cambia durante el desarrollo y la activación de los linfocitos B. Antes de la maduración de estos últimos, cuando aún no se han expuesto a su antígeno, se conocen como linfocitos B vírgenes y sólo expresan el isotipo IgM en su forma anclada a la superficie celular. Los linfocitos comienzan a expresar tanto IgM como IgD cuando alcanzan la madurez y en ese momento están listos para responder a su antígeno. La activación de los linfocitos B sigue al encuentro y unión de éste con su antígeno, lo que estimula a la célula para que se divide y se diferencie en una célula productora de anticuerpos denominada plasmática. En esta forma activada, los linfocitos B comienzan a secretar anticuerpos en lugar de anclarlos a la membrana. Algunas células hijas de los linfocitos B activados sufren un cambio isotípico, un mecanismo que provoca que la producción de anticuerpos en las formas IgM o IgD se trasmute a los otros tipos, IgE, IgA o IgG, que desempeñan distintos papeles en el sistema inmunitario.
IgG : Elevadas en Mieloma por IgG, Sarcoidosis, Enfermedad Hepática crónica, enf. autoinmunes, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgG. Su valor normal es 700-1500 mg/dl.
IgM : Elevadas en : Mieloma por IgM, Enfermedad Hepática crónica, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgM. Su nivel normal es 60-300 mg/dl.
IgA: Elevada en Mieloma por IgA, Cirrosis Hepática, infecciones crónicas, artritis reumatoidea, LES. Disminuida en Inmunodeficiencias adquiridas, Mieloma no por IgA. Su valor normal es 60-400 mg/dl.
IgE: Elevada en enfermedades atópicas, asma, fiebre del heno, infecciones parasitarias, Mieloma por IgE. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgE. Su valor norma es 0.002-0.2 mg/dl
IgD: Elevada en Infecciones crónicas, Mieloma por IgD, enfermedades autoinmunes. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgD. Su valor normal es 0-14 mg/dl
88347 La inmunoglobulina G (IgG) es una de las cinco clases de anticuerpos humorales producidos por el organismo. Se trata de la inmunoglobulina predominante en los fluidos internos del cuerpo, como son la sangre, el líquido cefalorraquídeo y el liquido peritoneal (líquido presente en la cavidad abdominal). Esta proteína especializada es sintetizada por el organismo en respuesta a la invasión de bacterias, hongos y virus. Es la inmunoglobulina más abundante del suero, con una concentración de 600-1.800 mg por 100 mL.1 La IgG constituye el 80% de las inmunoglobulinas totales. La IgG es la única clase de inmunoglobulinas que atraviesa la placenta, transmitiendo la inmunidad de la madre al feto. Es la inmunoglobulina más pequeña, con un peso molecular de 150.000 Daltons, así puede pasar fácilmente del sistema circulatorio del cuerpo a los tejidos. La síntesis de IgG se controla principalmente por el estímulo de los antígenos. En el caso de animales axénicos (sin microbios), con niveles de IgG muy bajos, el nivel de IgG se eleva en cuanto se les traslada a un ambiente normal. Se sabe que no todas las Inmunoglobulinas de una misma clase tienen idéntica estructura, sin que dentro de las clases se pueden establecer subclases considerando la secuencia de aminoácidos de la región constante de las cadenas H y el diferente número y situación de los puentes disulfuro intercatenarios establecidos entre las cadenas pesadas. Así, la IgG humana se divide en cuatro subclases (IgG1, IgG2, IgG3 e IgG4) y la IgA e IgM en dos (IgA1 e IgA2; IgM1 e IgM2) respectivamente. La inmunoglobulina M (IgM) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo, constituyendo un 6% de la población presente en sangre. Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso,macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons), aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM. La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizardeterminados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección. La inmunoglobulina A (IgA) es la clase predominante de anticuerpo en las secreciones sero mucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica. (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 mL.
88347 Las respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos. La inmunoglobulina D (IgD) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo humano. Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma. Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina. La IgD es una proteína funcional importante, pero su papel no es bien conocido; se sugiere que es un receptor antigénico de membrana, que conduciría a la diferenciación linfocitaria y también que es un ligando para los receptores de IgD en la inmunoregulación de las células T ‘helper’. Es muy susceptible a la proteólisis. La IgD representa menos del 1% del total de inmunoglobulinas plasmáticas; la concentración en suero depende de la edad y de la herencia genética. En suero de pacientes con mieloma IgD se encuentran concentraciones de IgD muy elevadas.
88347 La IgE es una glicoproteína de aproximadamente 190 kDa. Como otros anticuerpos monoméricos, está compuesta de dos cadenas pesadas y dos ligeras, unidas por puentes disulfuro. Su cadena pesada consta de 5 dominios, uno variable y cuatro constantes de tipo ε. La mayor parte de la IgE se encuentra unida a FcεRI en la superficie de los mastocitos, eosinófilos y basófilos. El reconocimiento de un antígeno por la IgE desencadena complejas reacciones inmunitarias, entre las que pueden destacarse, por ejemplo, la desgranulación de los mastocitos, que liberan sustancias vasoactivas como la histamina, así como la intervención de los eosinófilos en la respuesta inflamatoria. Contiene de un 10 a un 12% en peso de glúcidos y su concentración en el suero es de 0,01 a 0,10 mg por 100 mL. La producción de IgE específica de antígenos no patogénicos, como los encontrados en fármacos, alimentos, y alérgenos estacionales, puede provocar reacciones alérgicas. La liberación del contenido de los gránulos de los mastocitos al reconocimiento de antígeno causa síntomas como la hinchazón de las vías aéreas, la rinitis, y la dermatitis. La IgE se fija a la superficie de membrana de las células cebadas o basofilas hasta por 2 semanas en una respuesta alérgica. El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. La síntesis de las cadenas ligeras y pesadas se regula por genes que se encuentran en cromosomas distintos. Esto fue puesto de manifiesto empleando técnicas de digestión enzimática del DNA de células B y posterior hibridación con sondas de DNA complementario (cDNA), mediante la técnica de Southern Blot . Mediante esta técnica, se pudo observar que usando un cDNA marcado radioactivamente como sonda correspondiente a parte de una cadena pesada, éste se unía a segmentos de DNA de líneas celulares de ratón que contenían el cromosoma 12. Por otra parte, empleando igualmente sondas de cDNA marcadas radioactivamente frente al DNA de cadenas ligeras de tipo k, éstas se unen a células que contienen el cromosoma 6, mientras que las sondas para DNA de cadenas ligeras l lo hacen alcromosoma 16. Cuando una persona es alérgica a una sustancia en particular, el sistema inmunitario cree, erróneamente, que está bajo una invasión antígenica por parásitos, y produce la IgE, en un intento de "proteger" el organismo; de esta manera, se inicia una cadena de acontecimientos que provocan los síntomas de la alergia. Si una persona sufre de asma producida por reacciones alérgicas, esta cadena de acontecimientos también derivará en síntomas de asma. La molécula de IgD tiene una larga región "bisagra" entre Fab y Fc, lo que proporciona a la molécula más susceptibilidad a la degradación proteolítica, así como una mayor flexibilidad que favorece la unión al antígeno, y presenta 3 dominios en la región constante.
88347 Otra característica importante de la formación de inmunoglobulinas es que, en la síntesis de cada una de las cadenas de inmunoglobulinas participan varios segmentos de genes de cuyas combinaciones resultan los diversos genes funcionales, responsables directos de la codificación de cada una de las cadenas de inmunoglobulinas. Esto explica las múltiples posibilidades de formación del gran número de inmunoglobulinas partiendo de un limitado material genético. Estos segmentos codifican por separado la parte variable, la parte constante y las partes por las que ambas regiones se unen, esto es, las regiones bisagra. Los segmentos que codifican la parte variable son de dos tipos, segmentos V y segmentos D, responsables de la variabilidad y diversidad de las inmunoglobulinas respectivamente. Le siguen los segmentos de unión, responsables de unir los fragmentos anteriores con los segmentos C que codifican para la parte constante. El número de segmentos responsables de la codificación de la parte constante es muy limitado en relación con el número de segmentos que codifican la parte variable de las inmunoglobulinas. En la síntesis de cadenas ligeras participan los segmentos V, J y C (es decir, no hay genes D para la cadena ligera). Así pues, en el proceso de recombinación de genes de cadenas ligeras se acopla un segmento V con un segmento J y el conjunto V/J se recombina con el segmento correspondiente a la parte constante C. El proceso de transcripción se hace de tal manera que el RNA mensajero contiene información secuenciada V, J y C. A su vez, un proceso similar, si bien esta vez en dos etapas, ocurre para la formación de una cadena pesada. En este caso participan los segmentos V, D, J, y C. Primero se produce la recombinación entre un segmento D y un segmento J. En la segunda fase, este conjunto D/J se recombina con un segmento V. El complejo V/D/J se puede por último recombinar con cada uno de los segmentos que codifican las regiones constantes, según la inmunoglobulina a formar. El proceso de recombinación con los genes C tiene lugar a nivel de RNA. Vamos a encontrar en este momento el siguiente orden: el segmento líder; un fragmento V/D/J reordenado y unido y a continuación cada uno de los genes que codifican para las regiones constantes (Cm; Cd; Ct, etc) separados entre sí por los correspondientes intrones y a su vez precedidos por sus respectivos promotores. Se piensa que el gen Cm es el situado en primer lugar en la secuencia de genes C, seguido de Cd. En células B en reposo, el único y largo trascrito primario de RNA va a contener la información del complejo V/D/J más la correspondiente a los genes Cm y Cd. Mediante mecanismos de procesamiento del RNA que analizaremos posteriormente, se va a proceder a la escisión en el trascrito primario de la información correspondiente a Cm o bien a Cd. Se da lugar de esta manera a una inmunoglobulina de tipo IgM o IgD, y este mecanismo es además el responsable de que en las células B en reposo se encuentre la expresión simultánea de ambas inmunoglobulinas.
88347 El calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. El calostro de todas las especies mamíferas contiene ingredientes inmunizantes contra las bacterias Escherichia coli, que pueden llegar a causar diarrea y vómito. Además también neutralizan toxinas, virus y las bacterias en el sistema linfático y circulatorio. Se ha llegado a comprobar, que en las 72 h posteriores al alumbramiento, la vaca elimina hasta 2 kg de inmunoglobulinas. Puesto que los anticuerpos se dan de forma libre en el torrente sanguíneo, se dice que son parte del sistema inmunitario humoral. Los anticuerpos circulantes son producidos por líneas clonales de linfocitos B que responden específicamente a un antígeno que puede ser un fragmento de proteína de la cápside viral, por ejemplo. Los anticuerpos contribuyen a la inmunidad de tres formas distintas: pueden impedir que los patógenos entren en las células o las dañen al unirse a ellas (neutralización). Pueden estimular la eliminación de un patógeno por losmacrófagos y otras células revistiendo al patógeno (opsonización) y pueden desencadenar la destrucción directa del patógeno estimulando otrasrespuestas inmunes como la vía del complemento (lisis). Activación del complemento Los anticuerpos que se unen a la superficie de los antígenos, por ejemplo, en una bacteria, atraen los primeros componentes de la cascada del complemento mediante su región Fc e inician la activación del sistema "clásico" del complemento. Esto acaba con la muerte de la bacteria de dos formas: Primero, la unión de las moléculas del complemento con el anticuerpo marca al microbio para la ingestión por los fagocitos en un proceso llamado opsonización. Estos fagocitos son atraídos por ciertas moléculas del complemento. En segundo lugar, algunos componentes del sistema del complemento forman un complejo de ataque a membrana para ayudar a los anticuerpos a matar a la bacteria por medio de lisis. Los anticuerpos más efectivos en la activación del Sistema del Complemento son los de tipo IgM y los de tipo IgG subclase 1 y 3 (IgG1 e IgG3). Activación de células efectoras
88347 Para combatir a los patógenos que se replican en el exterior de las células, los anticuerpos se unen a los patógenos para ensamblarlos juntos provocando su aglutinación. Puesto que un anticuerpo tiene al menos dos paratopos se puede unir a más de un antígeno acoplándose a epítopos idénticos portados en las superficies de esos antígenos. Revistiendo al patógeno, los anticuerpos estimulan las funciones efectoras contra éste en las células que reconocen la región Fc. Aquellas células que reconocen los patógenos revestidos tienen receptores del Fc que, como su nombre indica, interactúan con la región Fc de los anticuerpos IgA, IgG, e IgE. El acoplamiento de un anticuerpo particular con el receptor Fc de una determinada célula desencadena en ella una función efectora: los fagocitos realizarán lafagocitosis, las células cebadas y los neutrófilos producirán la degranulación, las células asesinas naturales liberarán citoquinas y moléculas citotóxicas que finalmente acabarán con la destrucción del microbio invasor. Los receptores Fc son específicos del isotipo, lo que da una mayor flexibilidad al sistema inmune, afectando solo al mecanismo inmune adecuado para los distintos patógenos. La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T. Activación dependiente de células T Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie. La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores para la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipo IgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes. Activación T-independiente Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes: • Tipo 1, o activación policlonal T-independiente; • Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.
La inmunidad humoral es el principal mecanismo de defensa contra los microorganismos extracelulares y sus toxinas, en el cual, los componentes del sistema inmunitario que atacan a los antígenos, no son las células directamente sino los anticuerpos1 secretados por activación antigénica.
Tipos
Respuesta humoral primaria
La cantidad de anticuerpo secretado por células plasmáticas y la clonación de estas mismas células la primera vez que entra en contacto el receptor con el antígeno encuentra su máximo aproximadamente a los 7 días de la primera infección (5-10 días). Habitualmente, la respuesta máxima de anticuerpos son del isotipo IgM, por encima de IgG,2 inducida por todo tipo de inmunógeno. La dosis necesaria para la inmunización generalmente debe ser relativamente alta, óptimamente con la presencia de adyuvantes para los antígenos proteicos.
Respuesta humoral secundaria Una infección repetida por un mismo antígeno activa los linfocitos de memoria creados como consecuencia de la respuesta humoral primaria. La respuesta, entonces, se inicia más rápidamente, al cabo de unos 3 días. Por su parte, la respuesta máxima de anticuerpos es mayor, con una intensidad de 100 a 1000 veces la respuesta primaria, y es principalmente del isotipo IgG (en ciertas situaciones de los isotipos IgA e IgE). También dura más tiempo, haciendo que su declive sea más lento. Es una respuesta inducida por antígenos proteicos y sólo son requeridas bajas dosis de antígenos infectantes, sin necesidad de adyuvantes. Además de reconocer a los antígenos, los anticuerpos pueden neutralizar la capacidad de infectar de los microorganismos y "marcar" a los patógenos para facilitar su eliminación. Los anticuerpos no sólo son específicos en cuanto al tipo de antígeno que reconocen, sino también en cuanto al tipo de respuesta que inducen. Algunos promueven la fagocitosis y otros dificultan la secreción de mediadores químicos que participan en la respuesta inflamatoria y en la activación del complemento.
88033 La antigenicidad es la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. El lugar o determinante antigénico es la parte limitada de la molécula de un antígeno, adoptada especialmente a ciertas partes del anticuerpo específico: condiciona la aptitud del antígeno a combinarse selectivamente con el anticuerpo correspondiente. Un antígeno tiene generalmente varias localizaciones pudiendo unirse cada una de ellas con un anticuerpo específico diferente. Concepto relacionado: receptor de reconocimiento o de membrana, sensibilización y fragmento
88033 Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos, receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los epítopos de las células T se presentan en la superficie de una célula presentadora de antígeno, concretamente a través de moléculas del MHC. Los epítopos de las células T presentados por moléculas MHC clase I son típicamente péptidos de entre 8 y 15 aminoácidos de longitud, mientras que las moléculas MHC clase II presentan péptidos más largos y las moléculas MHC no clásicas también presentan epítopos no peptídicos como los glucolípidos. Los epítopos pueden ser trazados usando microarrays de proteínas, y con las técnicas ELISPOT o ELISA. Los epítopos tienen a veces reactividad cruzada. Esta propiedad es explotada por el sistema inmunitario en la regulación por anticuerpos anti-idiotípicos. Si un anticuerpo se une a un epítopo de un antígeno, el paratopo puede convertir el epítopo para otro anticuerpo que entonces se unirá a él. Si este segundo anticuerpo es de clase IgM, su unión puede acentuar la respuesta inmune; si el segundo anticuerpo es de clase IgG, su unión puede disminuirla. Actualmente se están realizando investigaciones intensivas para diseñar herramientas fiables que predecirán epítopos en proteínas.
IgG : Elevadas en Mieloma por IgG, Sarcoidosis, Enfermedad Hepática crónica, enf. autoinmunes, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgG. Su valor normal es 700-1500 mg/dl.
IgM : Elevadas en : Mieloma por IgM, Enfermedad Hepática crónica, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgM. Su nivel normal es 60-300 mg/dl.
IgA: Elevada en Mieloma por IgA, Cirrosis Hepática, infecciones crónicas, artritis reumatoidea, LES. Disminuida en Inmunodeficiencias adquiridas, Mieloma no por IgA. Su valor normal es 60-400 mg/dl.
IgE: Elevada en enfermedades atópicas, asma, fiebre del heno, infecciones parasitarias, Mieloma por IgE. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgE. Su valor norma es 0.002-0.2 mg/dl
IgD: Elevada en Infecciones crónicas, Mieloma por IgD, enfermedades autoinmunes. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgD. Su valor normal es 0-14 mg/dl
La inmunoglobulina M (IgM) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo, constituyendo un 6% de la población presente en sangre. Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso,macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons), aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM. La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizardeterminados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
La inmunoglobulina A (IgA) es la clase predominante de anticuerpo en las secreciones sero mucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica. (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentracLas respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
88033 Las respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
88033 La inmunoglobulina D (IgD) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo humano. Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma. Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina. La IgD es una proteína funcional importante, pero su papel no es bien conocido; se
88033 La IgE es una glicoproteína de aproximadamente 190 kDa. Como otros anticuerpos monoméricos, está compuesta de dos cadenas pesadas y dos ligeras, unidas por puentes disulfuro. Su cadena pesada consta de 5 dominios, uno variable y cuatro constantes de tipo ε. La mayor parte de la IgE se encuentra unida a FcεRI en la superficie de los mastocitos, eosinófilos y basófilos. El reconocimiento de un antígeno por la IgE desencadena complejas reacciones inmunitarias, entre las que pueden destacarse, por ejemplo, la desgranulación de los mastocitos, que liberan sustancias vasoactivas como la histamina, así como la intervención de los eosinófilos en la respuesta inflamatoria. Contiene de un 10 a un 12% en peso de glúcidos y su concentración en el suero es de 0,01 a 0,10 mg por 100 mL. La producción de IgE específica de antígenos no patogénicos, como los encontrados en fármacos, alimentos, y alérgenos estacionales, puede provocar reacciones alérgicas. La liberación del contenido de los gránulos de los mastocitos al reconocimiento de antígeno causa síntomas como la hinchazón de las vías aéreas, la rinitis, y la dermatitis. La IgE se fija a la superficie de membrana de las células cebadas o basofilas hasta por 2 semanas en una respuesta alérgica.
El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. La síntesis de las cadenas ligeras y pesadas se regula por genes que se encuentran en cromosomas distintos. Esto fue puesto de manifiesto empleando técnicas de digestión enzimática del DNA de células B y posterior hibridación con sondas de DNA complementario (cDNA), mediante la técnica de Southern Blot . Mediante esta técnica, se pudo observar que usando un cDNA marcado radioactivamente como sonda correspondiente a parte de una cadena pesada, éste se unía a segmentos de DNA de líneas celulares de ratón que contenían el cromosoma 12. Por otra parte, empleando igualmente sondas de cDNA marcadas radioactivamente frente al DNA de cadenas ligeras de tipo k, éstas se unen a células que contienen el cromosoma 6, mientras que las sondas para DNA de cadenas ligeras l lo hacen alcromosoma 16. Cuando una persona es alérgica a una sustancia en particular, el sistema inmunitario cree, erróneamente, que está bajo una invasión antígenica por parásitos, y produce la IgE, en un intento de "proteger" el organismo; de esta manera, se inicia una cadena de acontecimientos que provocan los síntomas de la alergia. Si una persona sufre de asma producida por reacciones alérgicas, esta cadena de acontecimientos también derivará en síntomas de asma. La molécula de IgD tiene una larga región "bisagra" entre Fab y Fc, lo que proporciona a la molécula más susceptibilidad a la degradación proteolítica, así como una mayor flexibilidad que favorece la unión al antígeno, y presenta 3 dominios en la región
88033 Otra característica importante de la formación de inmunoglobulinas es que, en la síntesis de cada una de las cadenas de inmunoglobulinas participan varios segmentos de genes de cuyas combinaciones resultan los diversos genes funcionales, responsables directos de la codificación de cada una de las cadenas de inmunoglobulinas. Esto explica las múltiples posibilidades de formación del gran número de inmunoglobulinas partiendo de un limitado material genético. Estos segmentos codifican por separado la parte variable, la parte constante y las partes por las que ambas regiones se unen, esto es, las regiones bisagra. Los segmentos que codifican la parte variable son de dos tipos, segmentos V y segmentos D, responsables de la variabilidad y diversidad de las inmunoglobulinas respectivamente. Le siguen los segmentos de unión, responsables de unir los fragmentos anteriores con los segmentos C que codifican para la parte constante. El número de segmentos responsables de la codificación de la parte constante es muy limitado en relación con el número de segmentos que codifican la parte variable de las inmunoglobulinas. En la síntesis de cadenas ligeras participan los segmentos V, J y C (es decir, no hay genes D para la cadena ligera). Así pues, en el proceso de recombinación de genes de cadenas ligeras se acopla un segmento V con un segmento J y el conjunto V/J se recombina con el segmento correspondiente a la parte constante C. El proceso de transcripción se hace de tal manera que el RNA mensajero contiene información secuenciada V, J y C. A su vez, un proceso similar, si bien esta vez en dos etapas,
88033 Funciones de los anticuerpos Puesto que los anticuerpos se dan de forma libre en el torrente sanguíneo, se dice que son parte del sistema inmunitario humoral. Los anticuerpos circulantes son producidos por líneas clonales de linfocitos B que responden específicamente a un antígeno que puede ser un fragmento de proteína de la cápside viral, por ejemplo. Los anticuerpos contribuyen a la inmunidad de tres formas distintas: pueden impedir que los patógenos entren en las células o las dañen al unirse a ellas (neutralización). Pueden estimular la eliminación de un patógeno por losmacrófagos y otras células revistiendo al patógeno (opsonización) y pueden desencadenar la destrucción directa del patógeno estimulando otrasrespuestas inmunes como la vía del complemento (lisis). Activación del complemento Los anticuerpos que se unen a la superficie de los antígenos, por ejemplo, en una bacteria, atraen los primeros componentes de la cascada del complemento mediante su región Fc e inician la activación del sistema "clásico" del complemento. Esto acaba con la muerte de la bacteria de dos formas: Primero, la unión de las moléculas del complemento con el anticuerpo marca al microbio para la ingestión por los fagocitos en un proceso llamado opsonización. Estos fagocitos son atraídos por ciertas moléculas del complemento. En segundo lugar, algunos componentes del sistema del complemento forman un complejo de ataque a membrana para ayudar a los anticuerpos a matar a la bacteria por medio de lisis. Los anticuerpos más efectivos en la activación del Sistema del Complemento son los de tipo IgM y los de tipo IgG subclase 1 y 3 (IgG1 e IgG3). La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T. Activación dependiente de células T Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie. La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores
para la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipo IgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes. Activación T-independiente Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes: • Tipo 1, o activación policlonal T-independiente; • Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.
88033 Las respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
El calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. El calostro de todas las especies mamíferas contiene ingredientes inmunizantes contra las bacterias Escherichia coli, que pueden llegar a causar diarrea y vómito. Además también neutralizan toxinas, virus y las bacterias en el sistema linfático y circulatorio. Se ha llegado a comprobar, que en las 72 h
88033 Regiones Fab y Fc Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla. consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla. consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos Los anticuerpos pueden presentarse en distintas variedades conocidas
Antigenicidad Capacidad para dar lugar a la producción de anticuerpos. El grado de antigenicidad depende del tipo y cuantía de la sustancia concreta, de las circunstancias del huésped y de su grado de sensibilidad al antígeno y de su capacidad para producir anticuerpos antigenia, o antigénico (potencial). Poder de comportarse como un antígeno, es decir, de provocar la formación de anticuerpos. Para ciertos autores, la antigenicidad sería la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. Difiere así del poder inmunógeno.
Es la unidad más pequeña de un antígeno que puede unirse a un BCR o TCR específico, o a los anticuerpos secretados. Es decir, los epitopos son las regiones inmunológicamente activas de un antígeno. Pueden ser lineales (continuos) o conformacionales. El sistema inmunológico humano es capaz de reconocer un amplio rango de distintos epítopos. Los anticuerpos son proteínas que produce el sistema inmune que reaccionan con su antígeno meta (que generalmente son proteínas presentes en organismos que invaden o materiales extraños tales como proteínas alimentarias). Si pensamos que los anticuerpos son llaves que se corresponden a una cerradura meta ubicada en el antígeno, el epítopo sería la ranura que corresponde a una llave específica. El epítopo suele ser un parche de residuos de aminoácidos, los bloques que forman las proteínas. El epítopo es la ubicación en el antígeno en donde la molécula del antígeno atraca, como una llave que abre su cerradura.
La estructura de los anticuerpos está basado en la cadena peptídica, una secuencia de aminoácidos sucedidos uno tras otro y unidos por sus funciones amino y carboxilo. Esta gran cadena posee distintos niveles de organización: primario, secundario, terciario e incluso cuaternario. están formados por dos subunidades, las cuales a su vez están compuestas por otras dos subunidades, una cadena liviana y una cadena pesada. Ambas cadenas se subdividen en una región constante y en una región variable. Esta última es la responsable de la unión del anticuerpo con el antígeno mientras que la región constante puede tener distintas funciones dependiendo del dominio. Existen distintos tipos de cadenas pesadas (mu, epsilon, alfa, gamma, delta) y distintos tipos de cadenas livianas (kappa y lambda), los cuales se encuentran determinados por la secuencia de aminoácidos de las regiones constantes.Se ha podido determinar dos zonas dentro de las inmunoglobulinas, ambas generadas a base de experimentos con enzimas. Un primer fragmento es el fragmento Fab (que puede unirse a los antígenos) y un segundo fragmento es el Fc (que no puede unirse a los antígenos pero sí puede servir como ligando de receptores Fc).Además, dentro de la región variable existen zonas más variables y regiones más conservadoras. Las primeras se denominan CDR y son 3, CDR1 , CDR 2, CDR 3. Esta última es la más variable.
Continuacion de estructura de las inmunoglobulinas
Cadenas Ligeras.
Hay dos tipos de cadenas ligeras, estructuralmente diferentes, que se conocen como cadenas ligeras tipo kappa (k) y cadenas ligeras tipo lambda (l). La familia de genes que codifica para la cadena ligera k se localiza en el cromosoma 2 y los loci de los genes homólogos que codifican para la cadena l, en el cromosoma 22. En cada molécula de inmunoglobulina las dos cadenas ligeras son del mismo tipo, k o bien l, pero nunca existe una de cada tipo en la misma inmunoglobulina. Las cadenas ligeras están formadas por unos 200 aminoácidos con la particularidad de que existen dos puentes disulfuro que unen grupos de unos cincuenta aminoácidos. Concretamente la IgG1 posee 214 aminoácidos y su estructura secundaria y terciaria están determinadas por dos puentes disulfuro intracatenarios que unen los aminoácidos 23 con el 88 y 134 con el 193 . A su vez, estas cadenas ligeras tienen otro puente disulfuro intercatenario, por el cual cada una de ellas se une a una cadena pesada para constituir la unidad básica de las inmunoglobulinas. Este puente se encuentra en el último aminoácido (214) de la parte carboxílica para el tipo k y en el penúltimo para el tipo l.
Cadenas pesadas. Estas cadenas poseen unos cuatrocientos aminoácidos estableciéndose entre algunos de ellos puentes disulfuro (intracatenarios) que asocian unos 60 aminoácidos y que condicionan la estructura secundaria del polipéptido. Por ejemplo, las cadenas pesadas de la IgG1 poseen 440 aminoácidos y los puentes disulfuro unen el aminoácido 22 con el 96, el 144 con el 200, el 261 con el 321 y el 367 con el 425. Estas dos cadenas pesadas están unidas la una a la otra por puentes disulfuro intercatenarios, ya indicados anteriormente, y que pueden ser de uno a cinco dependiendo del tipo de inmunoglobulina. En estas cadenas pesadas, y a nivel de los puentes disulfuro intercatenarios, hay una zona de unos 15 aminoácidos, de gran flexibilidad debido a su estructura y constituye lo que se denomina zona bisagra por donde se deforma la molécula de inmunoglobulina cuando se produce la unión con el antígeno, facilitándose así su acoplamiento con éste. Los loci de los genes que codifican para la cadena pesada se localizan en el brazo largo del cromosoma 14.
La información genética que permite la existencia de un repertorio amplio de anticuerpos reside en diferentes cromosomas: los genes que codifican para las diferentes regiones e isotipos de cadenas pesadas se encuentran en el cromosoma 14, los que codifican para la cadena ligera kappa se encuentran en el cromosoma 2 y en el 22, los que codifican para la cadena lambda. La distribución de los antisuerpos varía entre la vida prenatal y postnatal temprana (predominio de anticuerpos IgG maternos) y el resto de la vida del ser humano (predominio de los anticuerpos IgM en la respuesta inmune primaria y predominio global de los IgG en suero y de los IgA en secreciones mucosas, dentro de la respuesta secundaria). IgG: Es la inmunoglobulina de mayor concentración en circulación sanguínea (70% del total de los anticuerpos). Es un monómero, donde la cadena pesada puede ser de 4 subclases distintas. Es el anticuerpo preponderante en la respuesta secundaria. Todas las subclases atraviesan la placenta y todas, excepto la IgG4 pueden activar el complemento. Su localización puede ser tanto intravascular como extravascular. IgM: Es un anticuerpo que circula en sangre como un pentámero de la unidad básica. La molécula posee además una cadena adicional, llamada J, que parece desempeñar un papel importante en la polimerización de las subunidades. La IgM es el anticuerpo predominante en las respuestas primarias. La IgM encontrada en la superficie de los linfocitos B (BCR) es monomérica. IgA: Supone el 15-20% del total de imunoglobulinas, pero la mayoría (> 80%) circulan en el ser humano como un monómero de la unidad básica. Sin embargo, se encuentra en gran cantidad en las secreciones del tracto respiratorio, genitourinario, saliva, lágrimas, calostro y leche materna. En estas secreciones adopta una estructura dimérica que, al igual que la IgM, posee también una cadena J. Para poder atravesar los epitelios se une a un receptor de las células epiteliales que en un momento dado (después del transporte a la porción apical de la célula) sufre un clivaje y queda enganchado al dímero. A este fragmento de receptor, o cadena adicionada, se le denomina componente o pieza secretora y confiere al anticuerpo mayor resistencia al ataque proteolítico. Existen dos subclases de IgA, llamadas IgA1 e IgA2 funcionalmente equivalentes. IgD: Posee estructura monomérica. Constituye menos del 1% del total de anticuerpos circulantes, pero está presente, al igual que la IgM monomérica, en la superficie de todos los linfocitos B maduros vírgenes (BCR). Aunque se desconoce su función precisa, se sospecha que participa en la activación y/o diferenciación del linfocito B después del reconocimiento antigénico, a partir del cual cesa su expresión en la membrana. IgE: Posee una estructura monomérica. Es el anticuerpo de más baja concentración en circulación, pero se halla fijado (a través de su porción Fc) en la superficie de células cebadas y basófilos tisulares. Es el responsable de los fenómenos de hipersensibilidad inmediata y parece desempeñar un papel importante en la respuesta inmune contra ciertos parásitos (helmintos). Los anticuerpos reconocen al antígeno en su forma nativa y su unión se produce a través de los epitopos o determinantes antigénicos, por un lado, y las porciones hipervariables de las regiones variables de los anticuerpos. Las fuerzas que mantienen la unión no son covalentes (puentes de hidrógeno, interacciones electrostáticas, interacciones hidrófobas y fuerzas de Van der Waals) y esta unión mantiene diferentes grados de afinidad y avidez.
IgA es la clase predominante de anticuerpo en las secreciones seromucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica.1 (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 ml
es una de las cinco clases de anticuerpos humorales producidos por el organismo. Se trata de la inmunoglobulina predominante en los fluidos internos del cuerpo, como son la sangre, el líquido cefalorraquídeo y el liquido peritoneal (líquido presente en la cavidad abdominal). Esta proteína especializada es sintetizada por el organismo en respuesta a la invasión de bacterias, hongos y virus. Es la inmunoglobulina más abundante del suero, con una concentración de 600-1.800 mg por 100 mL.1 La IgG constituye el 80% de las inmunoglobulinas totales. La IgG es la única clase de inmunoglobulinas que atraviesa la placenta, transmitiendo la inmunidad de la madre al feto de manera natural y pasiva. Es la inmunoglobulina más pequeña, con un peso molecular de 150.000 Daltons,1 así puede pasar fácilmente del sistema circulatorio del cuerpo a los tejidos. La síntesis de IgG se controla principalmente por el estímulo de los antígenos. En el caso de animales axénicos (sin microbios), con niveles de IgG muy bajos, el nivel de IgG se eleva en cuanto se les traslada a un ambiente normal. Hay veces que estas inmunoglobulinas no responden adecuadamente. Especialmente en casos de Anaplasmosis simple. Las cadenas H de la IgG son del isotipo γ, contiene un 2-3% en peso de glúcidos.
IgM constituyendo un 6% de la población presente en sangre. También se encuentra presente en elasmobranquios, teleósteos, anfibios, reptiles y aves, siendo uno de los anticuerpos más antiguos en la historia evolutiva1 . Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso, macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons),2 aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM. La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizar determinados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
es la clase predominante de anticuerpo en las secreciones seromucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica.1 (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 mL.
Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma. Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina. La IgD es una proteína funcional importante, pero su papel no es bien conocido; se sugiere que es un receptor antigénico de membrana, que conduciría a la diferenciación linfocitaria y también que es un ligando para los receptores de IgD en la inmunoregulación de las células T ‘helper’. Es muy susceptible a la proteólisis. La IgD representa menos del 1% del total de inmunoglobulinas plasmáticas; la concentración en suero depende de la edad y de la herencia genética. En suero de pacientes con mieloma IgD se encuentran concentraciones de IgD muy elevadas.
El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. Cada linfo B rearregla su ADN para un gen funcional de cadena H y uno de cadena L. La especificidad de Ac se determina por la secuencia de aa de los dominios V de sus cadenas H y L, donde se capta Ag. La secuencia de la región C es la misma para las cadenas H y L de un isotipo determinado y no tiene efecto en la especificidad. La familia de Ig tiene un número pequeño de dominios de C, unidos en varias combinaciones con una variedad casi ilimitada de secuencias de la región V. La información está dispersa a lo largo del ADN, en múltiples segmentos codificadores (exones), separados por regiones ADN no codificador (intrones), cuando el gene se transcribe al ARN, los intrones se eliminan y los exones se unen.
Genes para cadenas ligeras La información genética necesaria para producir cadenas k se encuentra en un solo locus (k) en el cromosoma 2. Las cadenas l, nacen de un complejo genético similar en el cromosoma 22; la unión de segmentos Vl y Jl se presentan de una manera idéntica a los segmento k. Las cadenas H se derivan de una sola región del cromosoma 14; cada región C de la cadena H está codificada por un grupo de varios exones cortos. La secuencia de la región C (CH) para cada uno de los nueve isotipos de la cadena pesada, se rearreglan a lo largo del cromosoma; sólo esta presente una copia de cada uno.
Calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. "Para el nacimiento del bebé se preparara uno de los alimentos más importantes de su vida, el calostro. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. 87945



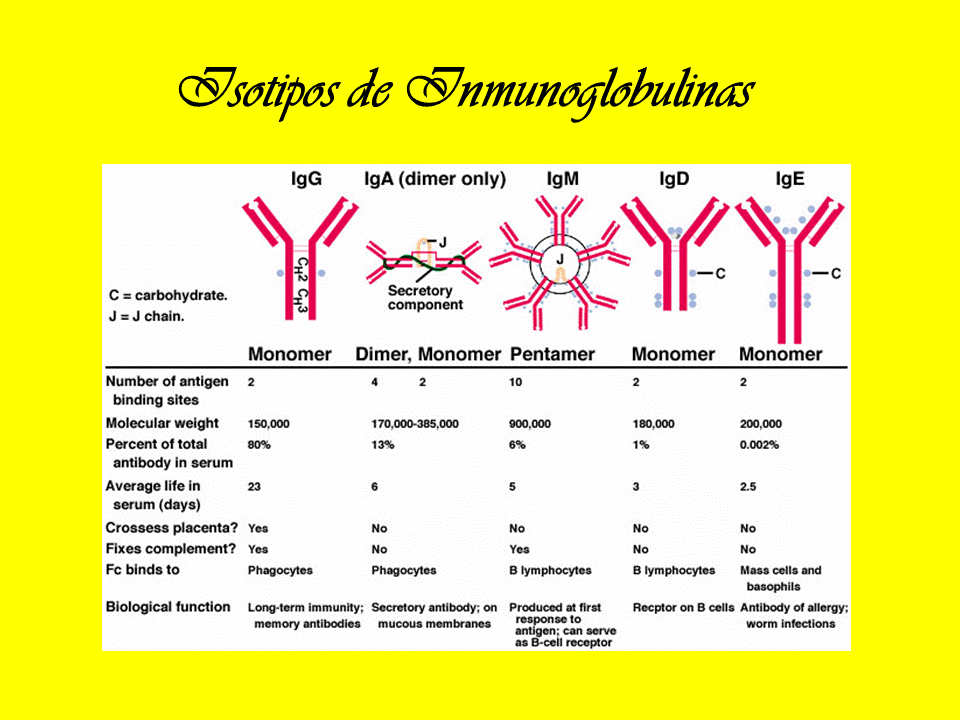













JAVIER DE LA TORRE 69412
ResponderEliminarLa inmunidad humoral es el principal mecanismo de defensa contra los microorganismos extracelulares y sus toxinas, en el cual, los componentes del sistema inmunitario que atacan a los antígenos, no son las células directamente sino los anticuerpos1 secretados por activación antigénica.
La respuesta efectora humoral está a cargo de las inmunoglobulinas o anticuerpos que son moléculas secretadas por células plasmáticas. Existen cinco clases de inmunoglobulinas: IgM, IgG, IgA, IgE e IgD, formadas por una unidad básica compuesta de dos cadenas polipetídicas globulares pesadas y dos cadenas livianas unidas entre sí por puentes disulfuro (A). Ambas cadenas presentan una zona constante (c) y una zona variable (v). En esta última, se encuentra una zona hipervariable formada por 10 a 15 aminoácidos que conforman el receptor idiotípico (r) responsable de la unión con el epitopo presente en el antígeno.
Al ser sometida a digestión por papaína (flecha), esta molécula genera dos fragmentos, el fragmento Fab responsable de la unión con antígeno y el fragmento Fc que determina diversas funciones biológicas en las diferentes inmunoglobulinas. El fragmento Fab está formado por una región constante y una región variable de una cadena pesada y de una cadena liviana. El fragmento Fc está formado sólo por regiones constantes de cadenas pesadas.
Las clases de inmunoglobulinas están determinadas por los diferentes isotipos de las cadenas pesadas. Estas pueden ser mu, gamma, alfa, delta o epsilon. Las cadenas livianas pueden ser kappa o bien lambda. Las distintas clases de inmunoglobulinas presentan diversas funciones biológicas.
JAVIER DE LA TORRE 69412
ResponderEliminaros Anticuerpos desempeñan un papel importante en el sistema inmune. Las inmunoglobulinas presentes en la superficie del B-Linfocito envían en señales a los electores citoplásmicos y nucleares. Éstos también entregan el antígeno a la célula donde puede ser destruido, ser tramitado y ser vuelto a la superficie de la célula que se presentará por las moléculas de la clase II de MHC a las células de ayudante del fi c T del antígeno-speci.
Los linfocitos de T a su vez envían señales a las células de B para que maduren y reconozcan los antígenos y creen los anticuerpos apuntados específicamente contra él.
Inmunorespuesta Humoral
Los Anticuerpos secretados por los linfocitos de B son responsables de la inmunorespuesta humoral. El sistema inmune humoral ayuda en la destrucción de patógeno externos y previene la extensión de infecciones intracelulares. Este sistema inmune también protege contra las toxinas.
Diversas funciones de diversas partes del anticuerpo
Las dos porciones estructurales del anticuerpo, del constante (Fc) los fragmentos es decir variables (Fabuloso) y, comunican funciones biológicas distintas.
michelle m cordero rodriguez 2011-0288
ResponderEliminarSlide #1
La antigenicidad es la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. El lugar o determinante antigénico es la parte limitada de la molécula de un antígeno, adoptada especialmente a ciertas partes del anticuerpo específico: condiciona la aptitud del antígeno a combinarse selectivamente con el anticuerpo correspondiente. Un antígeno tiene generalmente varias localizaciones pudiendo unirse cada una de ellas con un anticuerpo específico diferente. Concepto relacionado: receptor de reconocimiento o de membrana, sensibilización y fragmento Fab.
Slide #2
Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos, receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los epítopos de las células T se presentan en la superficie de una célula presentadora de antígeno, concretamente a través de moléculas del MHC. Los epítopos de las células T presentados por moléculas MHC clase I son típicamente péptidos de entre 8 y 15 aminoácidos de longitud, mientras que las moléculas MHC clase II presentan péptidos más largos y las moléculas MHC no clásicas también presentan epítopos no peptídicos como los glucolípidos. Los epítopos pueden ser trazados usando microarrays de proteínas, y con las técnicas ELISPOT o ELISA. Los epítopos tienen a veces reactividad cruzada. Esta propiedad es explotada por el sistema inmunitario en la regulación por anticuerpos anti-idiotípicos. Si un anticuerpo se une a un epítopo de un antígeno, el paratopo puede convertir el epítopo para otro anticuerpo que entonces se unirá a él. Si este segundo anticuerpo es de clase IgM, su unión puede acentuar la respuesta inmune; si el segundo anticuerpo es de clase IgG, su unión puede disminuirla. Actualmente se están realizando investigaciones intensivas para diseñar herramientas fiables que predecirán epítopos en proteínas.
michelle M. CORDERO rodriguez 2011-0288
EliminarSlide #3 Estructura de anticuerpo
Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ. El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos.
Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad. Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina. La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
Cadena ligera
En los mamíferos hay dos tipos de cadena ligera, llamados lambda (λ) y kappa (κ). Una cadena ligera contiene dos dominios sucesivos: un dominio constante y un dominio variable. La longitud aproximada de la cadena ligera es de 211 a 217 aminoácidos. Cada anticuerpo contiene dos cadenas ligeras que son siempre idénticas. Sólo un tipo de cadena ligera, κ o λ, está presente dentro del mismo anticuerpo en mamíferos. Otros tipos de cadenas ligeras como la cadena iota (ι), se encuentran en los vertebrados inferiores como los condrictios y teleósteos.
Regiones Fab y Fc
Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Slide #4 Tipos de anticuerpos
EliminarLos anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla.
El isotipo cambia durante el desarrollo y la activación de los linfocitos B. Antes de la maduración de estos últimos, cuando aún no se han expuesto a su antígeno, se conocen como linfocitos B vírgenes y sólo expresan el isotipo IgM en su forma anclada a la superficie celular. Los linfocitos comienzan a expresar tanto IgM como IgD cuando alcanzan la madurez y en ese momento están listos para responder a su antígeno. La activación de los linfocitos B sigue al encuentro y unión de éste con su antígeno, lo que estimula a la célula para que se divide y se diferencie en una célula productora de anticuerpos denominada plasmática. En esta forma activada, los linfocitos B comienzan a secretar anticuerpos en lugar de anclarlos a la membrana. Algunas células hijas de los linfocitos B activados sufren un cambio isotípico, un mecanismo que provoca que la producción de anticuerpos en las formas IgM o IgD se trasmute a los otros tipos, IgE, IgA o IgG, que desempeñan distintos papeles en el sistema inmunitario.
Slide #5 Inmunoglobulinas valores normales
IgG : Elevadas en Mieloma por IgG, Sarcoidosis, Enfermedad Hepática crónica, enf. autoinmunes, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgG. Su valor normal es 700-1500 mg/dl.
IgM : Elevadas en : Mieloma por IgM, Enfermedad Hepática crónica, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgM. Su nivel normal es 60-300 mg/dl.
IgA: Elevada en Mieloma por IgA, Cirrosis Hepática, infecciones crónicas, artritis reumatoidea, LES. Disminuida en Inmunodeficiencias adquiridas, Mieloma no por IgA. Su valor normal es 60-400 mg/dl.
IgE: Elevada en enfermedades atópicas, asma, fiebre del heno, infecciones parasitarias, Mieloma por IgE. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgE. Su valor norma es 0.002-0.2 mg/dl
IgD: Elevada en Infecciones crónicas, Mieloma por IgD, enfermedades autoinmunes. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgD. Su valor normal es 0-14 mg/dl
Slide #6 michelle m cordero 2011-0288
EliminarLa inmunoglobulina G (IgG) es una de las cinco clases de anticuerpos humorales producidos por el organismo. Se trata de la inmunoglobulina predominante en los fluidos internos del cuerpo, como son la sangre, el líquido cefalorraquídeo y el liquido peritoneal (líquido presente en la cavidad abdominal). Esta proteína especializada es sintetizada por el organismo en respuesta a la invasión de bacterias, hongos y virus. Es la inmunoglobulina más abundante del suero, con una concentración de 600-1.800 mg por 100 mL.1 La IgG constituye el 80% de las inmunoglobulinas totales. La IgG es la única clase de inmunoglobulinas que atraviesa la placenta, transmitiendo la inmunidad de la madre al feto. Es la inmunoglobulina más pequeña, con un peso molecular de 150.000 Daltons, así puede pasar fácilmente del sistema circulatorio del cuerpo a los tejidos. La síntesis de IgG se controla principalmente por el estímulo de los antígenos. En el caso de animales axénicos (sin microbios), con niveles de IgG muy bajos, el nivel de IgG se eleva en cuanto se les traslada a un ambiente normal. Se sabe que no todas las Inmunoglobulinas de una misma clase tienen idéntica estructura, sin que dentro de las clases se pueden establecer subclases considerando la secuencia de aminoácidos de la región constante de las cadenas H y el diferente número y situación de los puentes disulfuro intercatenarios establecidos entre las cadenas pesadas. Así, la IgG humana se divide en cuatro subclases (IgG1, IgG2, IgG3 e IgG4) y la IgA e IgM en dos (IgA1 e IgA2; IgM1 e IgM2) respectivamente.
Slide #7 IgM
La inmunoglobulina M (IgM) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo, constituyendo un 6% de la población presente en sangre. Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso,macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons), aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM.
La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizardeterminados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
Slide #8 michelle m cordero 2011-0288
EliminarLa inmunoglobulina A (IgA) es la clase predominante de anticuerpo en las secreciones sero mucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica. (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 mL.
Slide #9
Las respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
Slide #10
La inmunoglobulina D (IgD) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo humano. Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma.
Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina.
La IgD es una proteína funcional importante, pero su papel no es bien conocido; se sugiere que es un receptor antigénico de membrana, que conduciría a la diferenciación linfocitaria y también que es un ligando para los receptores de IgD en la inmunoregulación de las células T ‘helper’. Es muy susceptible a la proteólisis.
La IgD representa menos del 1% del total de inmunoglobulinas plasmáticas; la concentración en suero depende de la edad y de la herencia genética. En suero de pacientes con mieloma IgD se encuentran concentraciones de IgD muy elevadas.
Slide #11 michelle m cordero 2011-0288
ResponderEliminarLa IgE es una glicoproteína de aproximadamente 190 kDa. Como otros anticuerpos monoméricos, está compuesta de dos cadenas pesadas y dos ligeras, unidas por puentes disulfuro. Su cadena pesada consta de 5 dominios, uno variable y cuatro constantes de tipo ε. La mayor parte de la IgE se encuentra unida a FcεRI en la superficie de los mastocitos, eosinófilos y basófilos. El reconocimiento de un antígeno por la IgE desencadena complejas reacciones inmunitarias, entre las que pueden destacarse, por ejemplo, la desgranulación de los mastocitos, que liberan sustancias vasoactivas como la histamina, así como la intervención de los eosinófilos en la respuesta inflamatoria. Contiene de un 10 a un 12% en peso de glúcidos y su concentración en el suero es de 0,01 a 0,10 mg por 100 mL. La producción de IgE específica de antígenos no patogénicos, como los encontrados en fármacos, alimentos, y alérgenos estacionales, puede provocar reacciones alérgicas. La liberación del contenido de los gránulos de los mastocitos al reconocimiento de antígeno causa síntomas como la hinchazón de las vías aéreas, la rinitis, y la dermatitis. La IgE se fija a la superficie de membrana de las células cebadas o basofilas hasta por 2 semanas en una respuesta alérgica.
Slide 12
El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. La síntesis de las cadenas ligeras y pesadas se regula por genes que se encuentran en cromosomas distintos. Esto fue puesto de manifiesto empleando técnicas de digestión enzimática del DNA de células B y posterior hibridación con sondas de DNA complementario (cDNA), mediante la técnica de Southern Blot . Mediante esta técnica, se pudo observar que usando un cDNA marcado radioactivamente como sonda correspondiente a parte de una cadena pesada, éste se unía a segmentos de DNA de líneas celulares de ratón que contenían el cromosoma 12. Por otra parte, empleando igualmente sondas de cDNA marcadas radioactivamente frente al DNA de cadenas ligeras de tipo k, éstas se unen a células que contienen el cromosoma 6, mientras que las sondas para DNA de cadenas ligeras l lo hacen alcromosoma 16. Cuando una persona es alérgica a una sustancia en particular, el sistema inmunitario cree, erróneamente, que está bajo una invasión antígenica por parásitos, y produce la IgE, en un intento de "proteger" el organismo; de esta manera, se inicia una cadena de acontecimientos que provocan los síntomas de la alergia. Si una persona sufre de asma producida por reacciones alérgicas, esta cadena de acontecimientos también derivará en síntomas de asma. La molécula de IgD tiene una larga región "bisagra" entre Fab y Fc, lo que proporciona a la molécula más susceptibilidad a la degradación proteolítica, así como una mayor flexibilidad que favorece la unión al antígeno, y presenta 3 dominios en la región constante.
Slide #13
ResponderEliminarOtra característica importante de la formación de inmunoglobulinas es que, en la síntesis de cada una de las cadenas de inmunoglobulinas participan varios segmentos de genes de cuyas combinaciones resultan los diversos genes funcionales, responsables directos de la codificación de cada una de las cadenas de inmunoglobulinas. Esto explica las múltiples posibilidades de formación del gran número de inmunoglobulinas partiendo de un limitado material genético. Estos segmentos codifican por separado la parte variable, la parte constante y las partes por las que ambas regiones se unen, esto es, las regiones bisagra. Los segmentos que codifican la parte variable son de dos tipos, segmentos V y segmentos D, responsables de la variabilidad y diversidad de las inmunoglobulinas respectivamente. Le siguen los segmentos de unión, responsables de unir los fragmentos anteriores con los segmentos C que codifican para la parte constante. El número de segmentos responsables de la codificación de la parte constante es muy limitado en relación con el número de segmentos que codifican la parte variable de las inmunoglobulinas.
Slide 14
En la síntesis de cadenas ligeras participan los segmentos V, J y C (es decir, no hay genes D para la cadena ligera). Así pues, en el proceso de recombinación de genes de cadenas ligeras se acopla un segmento V con un segmento J y el conjunto V/J se recombina con el segmento correspondiente a la parte constante C. El proceso de transcripción se hace de tal manera que el RNA mensajero contiene información secuenciada V, J y C. A su vez, un proceso similar, si bien esta vez en dos etapas, ocurre para la formación de una cadena pesada. En este caso participan los segmentos V, D, J, y C. Primero se produce la recombinación entre un segmento D y un segmento J. En la segunda fase, este conjunto D/J se recombina con un segmento V. El complejo V/D/J se puede por último recombinar con cada uno de los segmentos que codifican las regiones constantes, según la inmunoglobulina a formar. El proceso de recombinación con los genes C tiene lugar a nivel de RNA. Vamos a encontrar en este momento el siguiente orden: el segmento líder; un fragmento V/D/J reordenado y unido y a continuación cada uno de los genes que codifican para las regiones constantes (Cm; Cd; Ct, etc) separados entre sí por los correspondientes intrones y a su vez precedidos por sus respectivos promotores. Se piensa que el gen Cm es el situado en primer lugar en la secuencia de genes C, seguido de Cd. En células B en reposo, el único y largo trascrito primario de RNA va a contener la información del complejo V/D/J más la correspondiente a los genes Cm y Cd. Mediante mecanismos de procesamiento del RNA que analizaremos posteriormente, se va a proceder a la escisión en el trascrito primario de la información correspondiente a Cm o bien a Cd. Se da lugar de esta manera a una inmunoglobulina de tipo IgM o IgD, y este mecanismo es además el responsable de que en las células B en reposo se encuentre la expresión simultánea de ambas inmunoglobulinas.
Slide #15 michelle m cordero 2011-0288
ResponderEliminarEl calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. El calostro de todas las especies mamíferas contiene ingredientes inmunizantes contra las bacterias Escherichia coli, que pueden llegar a causar diarrea y vómito. Además también neutralizan toxinas, virus y las bacterias en el sistema linfático y circulatorio. Se ha llegado a comprobar, que en las 72 h posteriores al alumbramiento, la vaca elimina hasta 2 kg de inmunoglobulinas.
michelle m. cordero 2011-0288 Slide# 16 Funciones de los Anticuerpos
ResponderEliminarPuesto que los anticuerpos se dan de forma libre en el torrente sanguíneo, se dice que son parte del sistema inmunitario humoral. Los anticuerpos circulantes son producidos por líneas clonales de linfocitos B que responden específicamente a un antígeno que puede ser un fragmento de proteína de la cápside viral, por ejemplo. Los anticuerpos contribuyen a la inmunidad de tres formas distintas: pueden impedir que los patógenos entren en las células o las dañen al unirse a ellas (neutralización). Pueden estimular la eliminación de un patógeno por losmacrófagos y otras células revistiendo al patógeno (opsonización) y pueden desencadenar la destrucción directa del patógeno estimulando otrasrespuestas inmunes como la vía del complemento (lisis).
Activación del complemento
Los anticuerpos que se unen a la superficie de los antígenos, por ejemplo, en una bacteria, atraen los primeros componentes de la cascada del complemento mediante su región Fc e inician la activación del sistema "clásico" del complemento. Esto acaba con la muerte de la bacteria de dos formas: Primero, la unión de las moléculas del complemento con el anticuerpo marca al microbio para la ingestión por los fagocitos en un proceso llamado opsonización. Estos fagocitos son atraídos por ciertas moléculas del complemento. En segundo lugar, algunos componentes del sistema del complemento forman un complejo de ataque a membrana para ayudar a los anticuerpos a matar a la bacteria por medio de lisis. Los anticuerpos más efectivos en la activación del Sistema del Complemento son los de tipo IgM y los de tipo IgG subclase 1 y 3 (IgG1 e IgG3).
michelle m cordero 2011-0288
ResponderEliminarActivación de células efectoras
Para combatir a los patógenos que se replican en el exterior de las células, los anticuerpos se unen a los patógenos para ensamblarlos juntos provocando su aglutinación. Puesto que un anticuerpo tiene al menos dos paratopos se puede unir a más de un antígeno acoplándose a epítopos idénticos portados en las superficies de esos antígenos. Revistiendo al patógeno, los anticuerpos estimulan las funciones efectoras contra éste en las células que reconocen la región Fc.
Aquellas células que reconocen los patógenos revestidos tienen receptores del Fc que, como su nombre indica, interactúan con la región Fc de los anticuerpos IgA, IgG, e IgE. El acoplamiento de un anticuerpo particular con el receptor Fc de una determinada célula desencadena en ella una función efectora: los fagocitos realizarán lafagocitosis, las células cebadas y los neutrófilos producirán la degranulación, las células asesinas naturales liberarán citoquinas y moléculas citotóxicas que finalmente acabarán con la destrucción del microbio invasor. Los receptores Fc son específicos del isotipo, lo que da una mayor flexibilidad al sistema inmune, afectando solo al mecanismo inmune adecuado para los distintos patógenos.
Slide# 17 Activación células B
EliminarLa activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T.
Activación dependiente de células T
Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie.
La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores
michelle m cordero 2011-0288
Eliminarpara la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipo IgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes.
Activación T-independiente
Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes:
• Tipo 1, o activación policlonal T-independiente;
• Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.
jose morales 86555
ResponderEliminarLa Inmunidad Humoral es aquella que se caracteriza porque los principales componentes del sistema Inmunológico, los Anticuerpos o Inmunoglobulinas bloquean o lisan Noxas biológicas en forma directa en el medio extracelular, es decir, sin haber ingresado a las células eucariotas del propio cuerpo, no hay participación directa de células Inmunológicas(Leucocitos-Linfocitos) sino solo los Anticuerpos sintetizados por los Linfocitos B y T.
En cambio la Inmunidad Celular es aquella en la cual participan directamente los Linfocitos T, bloqueando la proliferación de las Noxas Biológicas en el interior de las propias células eucariotas del cuerpo infectadas por Virus y Bacterias a la cual no tienen ecceso directo los Anticuerpos.
Se destacan los Linfocitos T Citotóxicos o Citolíticos que vuelcan en la membrana plasmática de las células del cuerpo infectadas Linfocinas, Citoquininas produciendo la Lisis de la propia célula infectada dejando en libertad a las Noxas para ser fagocitadas por otras células inmunológicas(Neutrófilos-Monocitos), las células T Cooperadoras que ayudan a los Linfocitos B y T Citotóxicos y las células Supresoras que se especializan en la activación o inhibición de las células B y T, aumentando su proliferación, Inhibiendo su proliferación o ambos procesos simultáneamente.
ALEJANDRA FIGUEROA 88099
ResponderEliminarCapacidad para dar lugar a la producción de anticuerpos. El grado de antigenicidad depende del tipo y cuantía de la sustancia concreta, de las circunstancias del huésped y de su grado de sensibilidad al antígeno y de su capacidad para producir anticuerpos.
Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos,1 receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos.
Los anticuerpos son proteínas plasmáticas globulares pesadas (~150 kDa), también conocidas como inmunoglobulinas. Tienen cadenas de azúcares unidas a alguno de sus residuos aminoácido.34 En otras palabras, los anticuerpos son glicoproteínas. La unidad básica funcional de cada anticuerpo es el monómero de inmunoglobulina, que contiene una sola unidad de Ig. Los anticuerpos secretados también pueden ser diméricos con dos unidades Ig, como en el caso de las IgA, tetraméricos con cuatro unidades Ig como en el caso de las IgM de teleósteo, o pentaméricos con cinco unidades de IgM, como en el caso de las IgM de mamíferos.
El monómero de Ig es una molécula en forma de "Y" que consta de dos cadenas de polipéptido; dos cadenas pesadas idénticas y dos cadenas ligeras idénticas conectadas por enlaces disulfuro.31 Cada cadena se compone de dominios estructurales llamados dominios Ig. Estos dominios contienen entre 70 y 110 aminoácidos y se clasifican en diferentes categorías, por ejemplo en variables (IgV) y constantes (IgC) de acuerdo con su tamaño y función.38 Tienen un "pliegue inmunoglobulina" característico en el cual dos láminas beta generan una forma de "sándwich", permaneciendo juntas por interacciones entre cisteínas bien conservadas a lo largo de la evolución, así como otros aminoácidos cargados.
Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ.3 El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos.3
1. Región Fab
2. Región Fc
3. Cadena pesada con un dominio variable (VH) seguido por un dominio constante (CH1), una región bisagra, y dos más constantes, los dominios (CH2 y CH3).
4. Cadena ligera con un dominio variable (VL) y uno constante (CL)
5. Lugar de unión al antígeno (paratopo)
6. Regiones bisagra.
Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad.31 Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina.3 La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
ALEJANDRA FIGUEROA 88099
EliminarLos anticuerpos (también conocidos como inmunoglobulinas, abreviado Ig) son glicoproteínas del tipo gamma globulina. Pueden encontrarse de forma soluble en la sangre u otros fluidos corporales de los vertebrados, disponiendo de una forma idéntica que actúa como receptor de los linfocitos B y son empleados por el sistema inmunitario para identificar y neutralizar elementos extraños tales como bacterias, virus o parásitos.1
El anticuerpo típico está constituido por unidades estructurales básicas, cada una de ellas con dos grandes cadenas pesadas y dos cadenas ligeras de menor tamaño, que forman, por ejemplo, monómeros con una unidad, dímeros con dos unidades o pentámeros con cinco unidades. Los anticuerpos son sintetizados por un tipo de leucocito denominado linfocito B. Existen distintas modalidades de anticuerpo, isotipos, basadas en la forma de cadena pesada que posean. Se conocen cinco clases diferentes de isotipos en mamíferos que desempeñan funciones diferentes, contribuyendo a dirigir la respuesta inmune adecuada para cada distinto tipo de cuerpo extraño que encuentran
Aunque la estructura general de todos los anticuerpos es muy semejante, una pequeña región del ápice de la proteína es extremadamente variable, lo cual permite la existencia de millones de anticuerpos, cada uno con un extremo ligeramente distinto. A esta parte de la proteína se la conoce como región hipervariable. Cada una de estas variantes se puede unir a una "diana" distinta, que es lo que se conoce como antígeno.3 Esta enorme diversidad de anticuerpos permite al sistema inmune reconocer una diversidad igualmente elevada de antígenos. La única parte del antígeno reconocida por el anticuerpo se denomina epítopo. Estos epítopos se unen con su anticuerpo en una interacción altamente específica que se denomina adaptación inducida, que permite a los anticuerpos identificar y unirse solamente a su antígeno único en medio de los millones de moléculas diferentes que componen un organismo.
El reconocimiento de un antígeno por un anticuerpo lo marca para ser atacado por otras partes del sistema inmunitario. Los anticuerpos también pueden neutralizar sus objetivos directamente, mediante, por ejemplo, la unión a una porción de un patógeno necesaria para que éste provoque una infección.
ALEJANDRA FIGUEROA 88099
EliminarLos linfocitos B son los leucocitos de los cuales depende la inmunidad mediada por anticuerpos con actividad específica de fijación de antígenos. Las células B, que constituyen un 5 a 15% del total de linfocitos,[cita requerida] dan origen a las células plasmáticas que producen anticuerpos.
Los linfocitos son de dos tipos principales, atendiendo a su origen y función: células T, que se diferencian inicialmente en el timo, y las células B, que se diferencian en el hígado y bazo fetal, y en la médula ósea del adulto (la 'B' proviene del latín Bursa fabricii, el órgano en el cual se desarrollan los linfocitos B en las aves). Durante su desarrollo, los linfocitos T y B adquieren receptores específicos para antígenos, el de las células B se le conoce como receptor de linfocito B (BCR).
La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T.
El calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo.
"Para el nacimiento del bebé se preparara uno de los alimentos más importantes de su vida, el calostro. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva.
El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros.
Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro.
Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé.
Dariana Japa 85113
ResponderEliminarHay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ. El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos.
Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad. Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina. La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
Cadena ligera
En los mamíferos hay dos tipos de cadena ligera, llamados lambda (λ) y kappa (κ). Una cadena ligera contiene dos dominios sucesivos: un dominio constante y un dominio variable. La longitud aproximada de la cadena ligera es de 211 a 217 aminoácidos. Cada anticuerpo contiene dos cadenas ligeras que son siempre idénticas. Sólo un tipo de cadena ligera, κ o λ, está presente dentro del mismo anticuerpo en mamíferos. Otros tipos de cadenas ligeras como la cadena iota (ι), se encuentran en los vertebrados inferiores como los condrictios y teleósteos.
Regiones Fab y Fc
Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Joan Marie Rodríguez 2011-0209
ResponderEliminarRespuesta Inmune Humoral
La ausencia de este tipo de respuesta deja al individuo tan indefenso frente a toda clase de gérmenes patógenos y otras agresiones, que es incompatible con la vida si no se instaura a tiempo un tratamiento adecuado.
La respuesta inmune humoral es mediatizada por los linfocitos B, que como se ha dicho anteriormente reconocen al antigeno a través de las inmunoglobulinas de membrana. Sin embargo este estimulo no es suficiente para que se inicien los procesos de proliferación de estas células. Para ello es necesario que los linfocitos B además del estimulo antigénico reciban el estimulo de ciertas interleucinas.
El elemento efector final de la respuesta humoral son las inmunoglobulinas. El termino inmunoglobulina fue propuesto por Heberman para designar a todas las sustancias con capacidad de anticuerpo, esto es con capacidad de anteponerse al antigeno. Las inmunoglobulinas son de cinco clases: inmunoglobulina M (IgM), inmunoglobulina A (IgA), inmunoglobulina G (IgG), inmunoglobulina D (IgD) e inmunoglobulina E (IgE). Las inmunoglobulinas tienen la propiedad de unirse específicamente al antígeno que indujo su formación.
Tras la unión antígeno-anticuerpo (Ag-Ac), las sustancias extrañas (o antígenos) son destruídas por las inmunoglobulinas a través de mecanismos, que pueden ser diferentes según el tipo de inmunoglobulina que participa. Esto se debe a que aunque las distintas clases de inmunoglobulinas tienen una estrucutura igual en ciertas partes de la molecula, en otras partes presentan una estrucura distinta. Podemos decir que las inmunoglobulinas, al detectar al antígeno y unirse a el, actúan como transductores de la información de la presencia de los mismos, que serán posteriormente destruidos por el mecanismo más idoneo, en el que colaborarán además del propio anticuerpo el sistema del complemento, macrófagos, los polimorfonucleares o células K.
El término complemento engloba, una gran variedad de proteinas, que interactúan en un determinado orden, se representan por C' y se encuentran en el suero. Cuando se produce la activación del C' se pone en marcha una serie de reacciones, en forma de "cascada", de tal forma que se van generando productos activos que además de influir en que la reacción prosiga tienen diferentes acciones biológicas importantes en la defensa del organismo.
Este comentario ha sido eliminado por un administrador del blog.
EliminarEste comentario ha sido eliminado por un administrador del blog.
EliminarEste comentario ha sido eliminado por un administrador del blog.
EliminarEste comentario ha sido eliminado por un administrador del blog.
EliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
EliminarEste comentario ha sido eliminado por un administrador del blog.
EliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminar88347
ResponderEliminarLa antigenicidad es la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. El lugar o determinante antigénico es la parte limitada de la molécula de un antígeno, adoptada especialmente a ciertas partes del anticuerpo específico: condiciona la aptitud del antígeno a combinarse selectivamente con el anticuerpo correspondiente. Un antígeno tiene generalmente varias localizaciones pudiendo unirse cada una de ellas con un anticuerpo específico diferente. Concepto relacionado: receptor de reconocimiento o de membrana, sensibilización y fragmento Fab.
Un epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos, receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los epítopos de las células T se presentan en la superficie de una célula presentadora de antígeno, concretamente a través de moléculas del MHC. Los epítopos de las células T presentados por moléculas MHC clase I son típicamente péptidos de entre 8 y 15 aminoácidos de longitud, mientras que las moléculas MHC clase II presentan péptidos más largos y las moléculas MHC no clásicas también presentan epítopos no peptídicos como los glucolípidos. Los epítopos pueden ser trazados usando microarrays de proteínas, y con las técnicas ELISPOT o ELISA. Los epítopos tienen a veces reactividad cruzada. Esta propiedad es explotada por el sistema inmunitario en la regulación por anticuerpos anti-idiotípicos. Si un anticuerpo se une a un epítopo de un antígeno, el paratopo puede convertir el epítopo para otro anticuerpo que entonces se unirá a él. Si este segundo anticuerpo es de clase IgM, su unión puede acentuar la respuesta inmune; si el segundo anticuerpo es de clase IgG, su unión puede disminuirla. Actualmente se están realizando investigaciones intensivas para diseñar herramientas fiables que predecirán epítopos en proteínas.
Hay cinco tipos de Ig en mamíferos que se nombran por letras griegas: α, δ, ε, γ y μ. El tipo de cadena pesada presente define la clase del anticuerpo. Estas cadenas se encuentran en los anticuerpos IgA, IgD, IgE, IgG, e IgM respectivamente. Las distintas cadenas pesadas difieren en tamaño y composición: α y γ contienen aproximadamente 450 aminoácidos, mientras que μ y ε poseen aproximadamente 550 aminoácidos.
Las cadenas pesadas γ, α y δ tienen una región constante compuesta de tres dominios estructurales Ig en tándem y una región bisagra para proporcionarle flexibilidad. Las cadenas pesadas μ y ε tienen una región constante compuesta por cuatro dominios inmunoglobulina. La región variable de la cadena pesada difiere en los anticuerpos producidos en los diferentes linfocitos B, pero es lo mismo para todos los anticuerpos producidos por el mismo linfocito B o por su línea clonal. La región variable de cada cadena pesada es de aproximadamente 110 aminoácidos y está compuesto por un único dominio Ig.
Cadena ligera
En los mamíferos hay dos tipos de cadena ligera, llamados lambda (λ) y kappa (κ). Una cadena ligera contiene dos dominios sucesivos: un dominio constante y un dominio variable. La longitud aproximada de la cadena ligera es de 211 a 217 aminoácidos. Cada anticuerpo contiene dos cadenas ligeras que son siempre idénticas. Sólo un tipo de cadena ligera, κ o λ, está presente dentro del mismo anticuerpo en mamíferos. Otros tipos de cadenas ligeras como la cadena iota (ι), se encuentran en los vertebrados inferiores como los condrictios y teleósteos.
88347
ResponderEliminarRegiones Fab y Fc
Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla.
El isotipo cambia durante el desarrollo y la activación de los linfocitos B. Antes de la maduración de estos últimos, cuando aún no se han expuesto a su antígeno, se conocen como linfocitos B vírgenes y sólo expresan el isotipo IgM en su forma anclada a la superficie celular. Los linfocitos comienzan a expresar tanto IgM como IgD cuando alcanzan la madurez y en ese momento están listos para responder a su antígeno. La activación de los linfocitos B sigue al encuentro y unión de éste con su antígeno, lo que estimula a la célula para que se divide y se diferencie en una célula productora de anticuerpos denominada plasmática. En esta forma activada, los linfocitos B comienzan a secretar anticuerpos en lugar de anclarlos a la membrana. Algunas células hijas de los linfocitos B activados sufren un cambio isotípico, un mecanismo que provoca que la producción de anticuerpos en las formas IgM o IgD se trasmute a los otros tipos, IgE, IgA o IgG, que desempeñan distintos papeles en el sistema inmunitario.
IgG : Elevadas en Mieloma por IgG, Sarcoidosis, Enfermedad Hepática crónica, enf. autoinmunes, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgG. Su valor normal es 700-1500 mg/dl.
IgM : Elevadas en : Mieloma por IgM, Enfermedad Hepática crónica, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgM. Su nivel normal es 60-300 mg/dl.
IgA: Elevada en Mieloma por IgA, Cirrosis Hepática, infecciones crónicas, artritis reumatoidea, LES. Disminuida en Inmunodeficiencias adquiridas, Mieloma no por IgA. Su valor normal es 60-400 mg/dl.
IgE: Elevada en enfermedades atópicas, asma, fiebre del heno, infecciones parasitarias, Mieloma por IgE. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgE. Su valor norma es 0.002-0.2 mg/dl
IgD: Elevada en Infecciones crónicas, Mieloma por IgD, enfermedades autoinmunes. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgD. Su valor normal es 0-14 mg/dl
88347
ResponderEliminarLa inmunoglobulina G (IgG) es una de las cinco clases de anticuerpos humorales producidos por el organismo. Se trata de la inmunoglobulina predominante en los fluidos internos del cuerpo, como son la sangre, el líquido cefalorraquídeo y el liquido peritoneal (líquido presente en la cavidad abdominal). Esta proteína especializada es sintetizada por el organismo en respuesta a la invasión de bacterias, hongos y virus. Es la inmunoglobulina más abundante del suero, con una concentración de 600-1.800 mg por 100 mL.1 La IgG constituye el 80% de las inmunoglobulinas totales. La IgG es la única clase de inmunoglobulinas que atraviesa la placenta, transmitiendo la inmunidad de la madre al feto. Es la inmunoglobulina más pequeña, con un peso molecular de 150.000 Daltons, así puede pasar fácilmente del sistema circulatorio del cuerpo a los tejidos. La síntesis de IgG se controla principalmente por el estímulo de los antígenos. En el caso de animales axénicos (sin microbios), con niveles de IgG muy bajos, el nivel de IgG se eleva en cuanto se les traslada a un ambiente normal. Se sabe que no todas las Inmunoglobulinas de una misma clase tienen idéntica estructura, sin que dentro de las clases se pueden establecer subclases considerando la secuencia de aminoácidos de la región constante de las cadenas H y el diferente número y situación de los puentes disulfuro intercatenarios establecidos entre las cadenas pesadas. Así, la IgG humana se divide en cuatro subclases (IgG1, IgG2, IgG3 e IgG4) y la IgA e IgM en dos (IgA1 e IgA2; IgM1 e IgM2) respectivamente.
La inmunoglobulina M (IgM) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo, constituyendo un 6% de la población presente en sangre. Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso,macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons), aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM.
La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizardeterminados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
La inmunoglobulina A (IgA) es la clase predominante de anticuerpo en las secreciones sero mucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica. (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 mL.
88347
ResponderEliminarLas respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
La inmunoglobulina D (IgD) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo humano. Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma.
Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina.
La IgD es una proteína funcional importante, pero su papel no es bien conocido; se sugiere que es un receptor antigénico de membrana, que conduciría a la diferenciación linfocitaria y también que es un ligando para los receptores de IgD en la inmunoregulación de las células T ‘helper’. Es muy susceptible a la proteólisis.
La IgD representa menos del 1% del total de inmunoglobulinas plasmáticas; la concentración en suero depende de la edad y de la herencia genética. En suero de pacientes con mieloma IgD se encuentran concentraciones de IgD muy elevadas.
88347
ResponderEliminarLa IgE es una glicoproteína de aproximadamente 190 kDa. Como otros anticuerpos monoméricos, está compuesta de dos cadenas pesadas y dos ligeras, unidas por puentes disulfuro. Su cadena pesada consta de 5 dominios, uno variable y cuatro constantes de tipo ε. La mayor parte de la IgE se encuentra unida a FcεRI en la superficie de los mastocitos, eosinófilos y basófilos. El reconocimiento de un antígeno por la IgE desencadena complejas reacciones inmunitarias, entre las que pueden destacarse, por ejemplo, la desgranulación de los mastocitos, que liberan sustancias vasoactivas como la histamina, así como la intervención de los eosinófilos en la respuesta inflamatoria. Contiene de un 10 a un 12% en peso de glúcidos y su concentración en el suero es de 0,01 a 0,10 mg por 100 mL. La producción de IgE específica de antígenos no patogénicos, como los encontrados en fármacos, alimentos, y alérgenos estacionales, puede provocar reacciones alérgicas. La liberación del contenido de los gránulos de los mastocitos al reconocimiento de antígeno causa síntomas como la hinchazón de las vías aéreas, la rinitis, y la dermatitis. La IgE se fija a la superficie de membrana de las células cebadas o basofilas hasta por 2 semanas en una respuesta alérgica.
El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. La síntesis de las cadenas ligeras y pesadas se regula por genes que se encuentran en cromosomas distintos. Esto fue puesto de manifiesto empleando técnicas de digestión enzimática del DNA de células B y posterior hibridación con sondas de DNA complementario (cDNA), mediante la técnica de Southern Blot . Mediante esta técnica, se pudo observar que usando un cDNA marcado radioactivamente como sonda correspondiente a parte de una cadena pesada, éste se unía a segmentos de DNA de líneas celulares de ratón que contenían el cromosoma 12. Por otra parte, empleando igualmente sondas de cDNA marcadas radioactivamente frente al DNA de cadenas ligeras de tipo k, éstas se unen a células que contienen el cromosoma 6, mientras que las sondas para DNA de cadenas ligeras l lo hacen alcromosoma 16. Cuando una persona es alérgica a una sustancia en particular, el sistema inmunitario cree, erróneamente, que está bajo una invasión antígenica por parásitos, y produce la IgE, en un intento de "proteger" el organismo; de esta manera, se inicia una cadena de acontecimientos que provocan los síntomas de la alergia. Si una persona sufre de asma producida por reacciones alérgicas, esta cadena de acontecimientos también derivará en síntomas de asma. La molécula de IgD tiene una larga región "bisagra" entre Fab y Fc, lo que proporciona a la molécula más susceptibilidad a la degradación proteolítica, así como una mayor flexibilidad que favorece la unión al antígeno, y presenta 3 dominios en la región constante.
88347
ResponderEliminarOtra característica importante de la formación de inmunoglobulinas es que, en la síntesis de cada una de las cadenas de inmunoglobulinas participan varios segmentos de genes de cuyas combinaciones resultan los diversos genes funcionales, responsables directos de la codificación de cada una de las cadenas de inmunoglobulinas. Esto explica las múltiples posibilidades de formación del gran número de inmunoglobulinas partiendo de un limitado material genético. Estos segmentos codifican por separado la parte variable, la parte constante y las partes por las que ambas regiones se unen, esto es, las regiones bisagra. Los segmentos que codifican la parte variable son de dos tipos, segmentos V y segmentos D, responsables de la variabilidad y diversidad de las inmunoglobulinas respectivamente. Le siguen los segmentos de unión, responsables de unir los fragmentos anteriores con los segmentos C que codifican para la parte constante. El número de segmentos responsables de la codificación de la parte constante es muy limitado en relación con el número de segmentos que codifican la parte variable de las inmunoglobulinas.
En la síntesis de cadenas ligeras participan los segmentos V, J y C (es decir, no hay genes D para la cadena ligera). Así pues, en el proceso de recombinación de genes de cadenas ligeras se acopla un segmento V con un segmento J y el conjunto V/J se recombina con el segmento correspondiente a la parte constante C. El proceso de transcripción se hace de tal manera que el RNA mensajero contiene información secuenciada V, J y C. A su vez, un proceso similar, si bien esta vez en dos etapas, ocurre para la formación de una cadena pesada. En este caso participan los segmentos V, D, J, y C. Primero se produce la recombinación entre un segmento D y un segmento J. En la segunda fase, este conjunto D/J se recombina con un segmento V. El complejo V/D/J se puede por último recombinar con cada uno de los segmentos que codifican las regiones constantes, según la inmunoglobulina a formar. El proceso de recombinación con los genes C tiene lugar a nivel de RNA. Vamos a encontrar en este momento el siguiente orden: el segmento líder; un fragmento V/D/J reordenado y unido y a continuación cada uno de los genes que codifican para las regiones constantes (Cm; Cd; Ct, etc) separados entre sí por los correspondientes intrones y a su vez precedidos por sus respectivos promotores. Se piensa que el gen Cm es el situado en primer lugar en la secuencia de genes C, seguido de Cd. En células B en reposo, el único y largo trascrito primario de RNA va a contener la información del complejo V/D/J más la correspondiente a los genes Cm y Cd. Mediante mecanismos de procesamiento del RNA que analizaremos posteriormente, se va a proceder a la escisión en el trascrito primario de la información correspondiente a Cm o bien a Cd. Se da lugar de esta manera a una inmunoglobulina de tipo IgM o IgD, y este mecanismo es además el responsable de que en las células B en reposo se encuentre la expresión simultánea de ambas inmunoglobulinas.
88347
ResponderEliminarEl calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. El calostro de todas las especies mamíferas contiene ingredientes inmunizantes contra las bacterias Escherichia coli, que pueden llegar a causar diarrea y vómito. Además también neutralizan toxinas, virus y las bacterias en el sistema linfático y circulatorio. Se ha llegado a comprobar, que en las 72 h posteriores al alumbramiento, la vaca elimina hasta 2 kg de inmunoglobulinas.
Puesto que los anticuerpos se dan de forma libre en el torrente sanguíneo, se dice que son parte del sistema inmunitario humoral. Los anticuerpos circulantes son producidos por líneas clonales de linfocitos B que responden específicamente a un antígeno que puede ser un fragmento de proteína de la cápside viral, por ejemplo. Los anticuerpos contribuyen a la inmunidad de tres formas distintas: pueden impedir que los patógenos entren en las células o las dañen al unirse a ellas (neutralización). Pueden estimular la eliminación de un patógeno por losmacrófagos y otras células revistiendo al patógeno (opsonización) y pueden desencadenar la destrucción directa del patógeno estimulando otrasrespuestas inmunes como la vía del complemento (lisis).
Activación del complemento
Los anticuerpos que se unen a la superficie de los antígenos, por ejemplo, en una bacteria, atraen los primeros componentes de la cascada del complemento mediante su región Fc e inician la activación del sistema "clásico" del complemento. Esto acaba con la muerte de la bacteria de dos formas: Primero, la unión de las moléculas del complemento con el anticuerpo marca al microbio para la ingestión por los fagocitos en un proceso llamado opsonización. Estos fagocitos son atraídos por ciertas moléculas del complemento. En segundo lugar, algunos componentes del sistema del complemento forman un complejo de ataque a membrana para ayudar a los anticuerpos a matar a la bacteria por medio de lisis. Los anticuerpos más efectivos en la activación del Sistema del Complemento son los de tipo IgM y los de tipo IgG subclase 1 y 3 (IgG1 e IgG3).
Activación de células efectoras
88347
ResponderEliminarPara combatir a los patógenos que se replican en el exterior de las células, los anticuerpos se unen a los patógenos para ensamblarlos juntos provocando su aglutinación. Puesto que un anticuerpo tiene al menos dos paratopos se puede unir a más de un antígeno acoplándose a epítopos idénticos portados en las superficies de esos antígenos. Revistiendo al patógeno, los anticuerpos estimulan las funciones efectoras contra éste en las células que reconocen la región Fc.
Aquellas células que reconocen los patógenos revestidos tienen receptores del Fc que, como su nombre indica, interactúan con la región Fc de los anticuerpos IgA, IgG, e IgE. El acoplamiento de un anticuerpo particular con el receptor Fc de una determinada célula desencadena en ella una función efectora: los fagocitos realizarán lafagocitosis, las células cebadas y los neutrófilos producirán la degranulación, las células asesinas naturales liberarán citoquinas y moléculas citotóxicas que finalmente acabarán con la destrucción del microbio invasor. Los receptores Fc son específicos del isotipo, lo que da una mayor flexibilidad al sistema inmune, afectando solo al mecanismo inmune adecuado para los distintos patógenos.
La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T.
Activación dependiente de células T
Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie.
La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores
para la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipo IgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes.
Activación T-independiente
Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes:
• Tipo 1, o activación policlonal T-independiente;
• Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.
Matricula 75596
ResponderEliminarInmunidad Humoral
La inmunidad humoral es el principal mecanismo de defensa contra los microorganismos extracelulares y sus toxinas, en el cual, los componentes del sistema inmunitario que atacan a los antígenos, no son las células directamente sino los anticuerpos1 secretados por activación antigénica.
Tipos
Respuesta humoral primaria
La cantidad de anticuerpo secretado por células plasmáticas y la clonación de estas mismas células la primera vez que entra en contacto el receptor con el antígeno encuentra su máximo aproximadamente a los 7 días de la primera infección (5-10 días). Habitualmente, la respuesta máxima de anticuerpos son del isotipo IgM, por encima de IgG,2 inducida por todo tipo de inmunógeno. La dosis necesaria para la inmunización generalmente debe ser relativamente alta, óptimamente con la presencia de adyuvantes para los antígenos proteicos.
Respuesta humoral secundaria
Una infección repetida por un mismo antígeno activa los linfocitos de memoria creados como consecuencia de la respuesta humoral primaria. La respuesta, entonces, se inicia más rápidamente, al cabo de unos 3 días. Por su parte, la respuesta máxima de anticuerpos es mayor, con una intensidad de 100 a 1000 veces la respuesta primaria, y es principalmente del isotipo IgG (en ciertas situaciones de los isotipos IgA e IgE). También dura más tiempo, haciendo que su declive sea más lento. Es una respuesta inducida por antígenos proteicos y sólo son requeridas bajas dosis de antígenos infectantes, sin necesidad de adyuvantes.
Además de reconocer a los antígenos, los anticuerpos pueden neutralizar la capacidad de infectar de los microorganismos y "marcar" a los patógenos para facilitar su eliminación.
Los anticuerpos no sólo son específicos en cuanto al tipo de antígeno que reconocen, sino también en cuanto al tipo de respuesta que inducen. Algunos promueven la fagocitosis y otros dificultan la secreción de mediadores químicos que participan en la respuesta inflamatoria y en la activación del complemento.
88033
ResponderEliminarLa antigenicidad es la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. El lugar o determinante antigénico es la parte limitada de la molécula de un antígeno, adoptada especialmente a ciertas partes del anticuerpo específico: condiciona la aptitud del antígeno a combinarse selectivamente con el anticuerpo correspondiente. Un antígeno tiene generalmente varias localizaciones pudiendo unirse cada una de ellas con un anticuerpo específico diferente. Concepto relacionado: receptor de reconocimiento o de membrana, sensibilización y fragmento
88033
ResponderEliminarUn epítopo o determinante antigénico es la porción de una macromolécula que es reconocida por el sistema inmunitario, específicamente la secuencia específica a la que se unen los anticuerpos, receptores de las células B o de células T. Aunque se piensa que los epítopos provienen de proteínas no propias, las secuencias que se obtienen del huésped que pueden ser reconocidas son también clasificadas como epítopos. Los epítopos de las células T se presentan en la superficie de una célula presentadora de antígeno, concretamente a través de moléculas del MHC. Los epítopos de las células T presentados por moléculas MHC clase I son típicamente péptidos de entre 8 y 15 aminoácidos de longitud, mientras que las moléculas MHC clase II presentan péptidos más largos y las moléculas MHC no clásicas también presentan epítopos no peptídicos como los glucolípidos. Los epítopos pueden ser trazados usando microarrays de proteínas, y con las técnicas ELISPOT o ELISA. Los epítopos tienen a veces reactividad cruzada. Esta propiedad es explotada por el sistema inmunitario en la regulación por anticuerpos anti-idiotípicos. Si un anticuerpo se une a un epítopo de un antígeno, el paratopo puede convertir el epítopo para otro anticuerpo que entonces se unirá a él. Si este segundo anticuerpo es de clase IgM, su unión puede acentuar la respuesta inmune; si el segundo anticuerpo es de clase IgG, su unión puede disminuirla. Actualmente se están realizando investigaciones intensivas para diseñar herramientas fiables que predecirán epítopos en proteínas.
88033
ResponderEliminarInmunoglobulinas valores normales
IgG : Elevadas en Mieloma por IgG, Sarcoidosis, Enfermedad Hepática crónica, enf. autoinmunes, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgG. Su valor normal es 700-1500 mg/dl.
IgM : Elevadas en : Mieloma por IgM, Enfermedad Hepática crónica, infecciones crónicas. Disminuida en Inmunodeficiencias adquiridas, defecto hereditario, embarazo, mieloma no por IgM. Su nivel normal es 60-300 mg/dl.
IgA: Elevada en Mieloma por IgA, Cirrosis Hepática, infecciones crónicas, artritis reumatoidea, LES. Disminuida en Inmunodeficiencias adquiridas, Mieloma no por IgA. Su valor normal es 60-400 mg/dl.
IgE: Elevada en enfermedades atópicas, asma, fiebre del heno, infecciones parasitarias, Mieloma por IgE. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgE. Su valor norma es 0.002-0.2 mg/dl
IgD: Elevada en Infecciones crónicas, Mieloma por IgD, enfermedades autoinmunes. Disminuida en déficits hereditarios, inmunodeficiencias adquiridas, Mieloma no por IgD. Su valor normal es 0-14 mg/dl
88033
ResponderEliminarLa inmunoglobulina M (IgM) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo, constituyendo un 6% de la población presente en sangre. Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso,macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons), aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM.
La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizardeterminados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
88033
ResponderEliminarLa inmunoglobulina A (IgA) es la clase predominante de anticuerpo en las secreciones sero mucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica. (IgA secretora) Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas. La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentracLas respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
88033
ResponderEliminarLas respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
88033
ResponderEliminarLa inmunoglobulina D (IgD) es uno de los cinco isotipos de inmunoglobulina (G, A, M, E, D) presentes en el organismo humano. Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma.
Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina.
La IgD es una proteína funcional importante, pero su papel no es bien conocido; se
88033
ResponderEliminarLa IgE es una glicoproteína de aproximadamente 190 kDa. Como otros anticuerpos monoméricos, está compuesta de dos cadenas pesadas y dos ligeras, unidas por puentes disulfuro. Su cadena pesada consta de 5 dominios, uno variable y cuatro constantes de tipo ε. La mayor parte de la IgE se encuentra unida a FcεRI en la superficie de los mastocitos, eosinófilos y basófilos. El reconocimiento de un antígeno por la IgE desencadena complejas reacciones inmunitarias, entre las que pueden destacarse, por ejemplo, la desgranulación de los mastocitos, que liberan sustancias vasoactivas como la histamina, así como la intervención de los eosinófilos en la respuesta inflamatoria. Contiene de un 10 a un 12% en peso de glúcidos y su concentración en el suero es de 0,01 a 0,10 mg por 100 mL. La producción de IgE específica de antígenos no patogénicos, como los encontrados en fármacos, alimentos, y alérgenos estacionales, puede provocar reacciones alérgicas. La liberación del contenido de los gránulos de los mastocitos al reconocimiento de antígeno causa síntomas como la hinchazón de las vías aéreas, la rinitis, y la dermatitis. La IgE se fija a la superficie de membrana de las células cebadas o basofilas hasta por 2 semanas en una respuesta alérgica.
88033
ResponderEliminarEl sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. La síntesis de las cadenas ligeras y pesadas se regula por genes que se encuentran en cromosomas distintos. Esto fue puesto de manifiesto empleando técnicas de digestión enzimática del DNA de células B y posterior hibridación con sondas de DNA complementario (cDNA), mediante la técnica de Southern Blot . Mediante esta técnica, se pudo observar que usando un cDNA marcado radioactivamente como sonda correspondiente a parte de una cadena pesada, éste se unía a segmentos de DNA de líneas celulares de ratón que contenían el cromosoma 12. Por otra parte, empleando igualmente sondas de cDNA marcadas radioactivamente frente al DNA de cadenas ligeras de tipo k, éstas se unen a células que contienen el cromosoma 6, mientras que las sondas para DNA de cadenas ligeras l lo hacen alcromosoma 16. Cuando una persona es alérgica a una sustancia en particular, el sistema inmunitario cree, erróneamente, que está bajo una invasión antígenica por parásitos, y produce la IgE, en un intento de "proteger" el organismo; de esta manera, se inicia una cadena de acontecimientos que provocan los síntomas de la alergia. Si una persona sufre de asma producida por reacciones alérgicas, esta cadena de acontecimientos también derivará en síntomas de asma. La molécula de IgD tiene una larga región "bisagra" entre Fab y Fc, lo que proporciona a la molécula más susceptibilidad a la degradación proteolítica, así como una mayor flexibilidad que favorece la unión al antígeno, y presenta 3 dominios en la región
88033
ResponderEliminarOtra característica importante de la formación de inmunoglobulinas es que, en la síntesis de cada una de las cadenas de inmunoglobulinas participan varios segmentos de genes de cuyas combinaciones resultan los diversos genes funcionales, responsables directos de la codificación de cada una de las cadenas de inmunoglobulinas. Esto explica las múltiples posibilidades de formación del gran número de inmunoglobulinas partiendo de un limitado material genético. Estos segmentos codifican por separado la parte variable, la parte constante y las partes por las que ambas regiones se unen, esto es, las regiones bisagra. Los segmentos que codifican la parte variable son de dos tipos, segmentos V y segmentos D, responsables de la variabilidad y diversidad de las inmunoglobulinas respectivamente. Le siguen los segmentos de unión, responsables de unir los fragmentos anteriores con los segmentos C que codifican para la parte constante. El número de segmentos responsables de la codificación de la parte constante es muy limitado en relación con el número de segmentos que codifican la parte variable de las
inmunoglobulinas.
En la síntesis de cadenas ligeras participan los segmentos V, J y C (es decir, no hay genes D para la cadena ligera). Así pues, en el proceso de recombinación de genes de cadenas ligeras se acopla un segmento V con un segmento J y el conjunto V/J se recombina con el segmento correspondiente a la parte constante C. El proceso de transcripción se hace de tal manera que el RNA mensajero contiene información secuenciada V, J y C. A su vez, un proceso similar, si bien esta vez en dos etapas,
88033
ResponderEliminarFunciones de los anticuerpos
Puesto que los anticuerpos se dan de forma libre en el torrente sanguíneo, se dice que son parte del sistema inmunitario humoral. Los anticuerpos circulantes son producidos por líneas clonales de linfocitos B que responden específicamente a un antígeno que puede ser un fragmento de proteína de la cápside viral, por ejemplo. Los anticuerpos contribuyen a la inmunidad de tres formas distintas: pueden impedir que los patógenos entren en las células o las dañen al unirse a ellas (neutralización). Pueden estimular la eliminación de un patógeno por losmacrófagos y otras células revistiendo al patógeno (opsonización) y pueden desencadenar la destrucción directa del patógeno estimulando otrasrespuestas inmunes como la vía del complemento (lisis).
Activación del complemento
Los anticuerpos que se unen a la superficie de los antígenos, por ejemplo, en una bacteria, atraen los primeros componentes de la cascada del complemento mediante su región Fc e inician la activación del sistema "clásico" del complemento. Esto acaba con la muerte de la bacteria de dos formas: Primero, la unión de las moléculas del complemento con el anticuerpo marca al microbio para la ingestión por los fagocitos en un proceso llamado opsonización. Estos fagocitos son atraídos por ciertas moléculas del complemento. En segundo lugar, algunos componentes del sistema del complemento forman un complejo de ataque a membrana para ayudar a los anticuerpos a matar a la bacteria por medio de lisis. Los anticuerpos más efectivos en la activación del Sistema del Complemento son los de tipo IgM y los de tipo IgG subclase 1 y 3 (IgG1 e IgG3).
La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T.
Activación dependiente de células T
Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie.
La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores
88033
ResponderEliminarpara la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipo IgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes.
Activación T-independiente
Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes:
• Tipo 1, o activación policlonal T-independiente;
• Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.
88033
ResponderEliminarLas respuestas inmunes humorales en la mucosa son primordialmente del isotipo IgA. La IgA secretora es un anticuerpo que puede atravesar las membranas mucosas y ayuda a impedir la entrada de los microorganismos. Un motivo para considerar el MALT como un sistema distinto, es que las células linfoides asociadas a la mucosa recirculan principalmente dentro del sistema linfoide mucoso. Así, las células linfoides estimuladas en las placas de Peyer por un antígeno, viajan por la linfa hacia los linfáticos regionales y luego por el conducto torácico a la circulación, para de nuevo ir a asentarse en las mucosas, donde van a secretar IgA. La recirculación específica (tropismo por las mucosas) se basa en las moléculas de adhesión que éstas células expresan. Basándose en este mecanismo, la estimulación antigénica en un área de la mucosa provoca una respuesta de anticuerpos (IgA) en otras zonas mucosas. Papel Biológico de la IgA de las Secreciones: 1) En la Luz tiene una función antiviral (tanto frente virus que producen infecciones locales como a virus que se diseminan). La IgAs neutraliza los virus impidiendo la colonización de la mucosa. También desarrolla una función antibacteriana porque impide la adherencia y colonización de las superficies mucosas. Además puede neutralizar toxinas bacterianas (cólera) y llevar a cabo una acción antiparasitaria. 2) Intraepitelial: Neutralización de los virus dentro de las células endoteliales infectadas. 3 ) En la Lámina Propia: Se forman muchos complejos inmunes dado el contacto tan frecuente con los antígenos. La IgAs contribuirá a eliminarlos hacia la luz (Ag + IgG + IgA). Por otro lado, la IgA es muy poco imflamatoria (activa poco al Complemento). Los complejos formados con IgA tienen muchas menos posibilidades de desencadenar enfermedad por inmunocomplejos.
88033
ResponderEliminarEl calostro es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva. El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros. Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro. Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé. El calostro de todas las especies mamíferas contiene ingredientes inmunizantes contra las bacterias Escherichia coli, que pueden llegar a causar diarrea y vómito. Además también neutralizan toxinas, virus y las bacterias en el sistema linfático y circulatorio. Se ha llegado a comprobar, que en las 72 h
88033
ResponderEliminarRegiones Fab y Fc
Algunas partes del anticuerpo tienen funciones únicas. Los extremos de la "Y", por ejemplo, contienen el lugar que se une al antígeno y por tanto, reconoce elementos
extraños específicos. Esta región del anticuerpo se llama Fragmento de unión al
antígeno o región Fab. Está compuesta de un dominio constante y otro variable de cada una de las cadenas ligera y pesada del anticuerpo. El paratopo está conformado por los dominios variables de la cadena pesada y ligera en el extremo
amino terminal del monómero de anticuerpo. El papel que desempeña la base de la "Y" consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla.
consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Los anticuerpos pueden presentarse en distintas variedades conocidas comoisotipos o clases. En mamíferos placentados existen cinco isotipos de anticuerpos conocidos como IgA, IgD, IgE,IgG e IgM. Se nombran mediante el prefijo "Ig" que significa inmunoglobulina y difieren en sus propiedades biológicas, localizaciones funcionales y capacidad para reconocer diferentes tipos de antígenos como se muestra en la tabla. consiste en modular la actividad de la célula inmunitaria. Esta región se llama Fragmento cristalizable o Fc y está compuesta por dos o tres dominios constantes de ambas cadenas pesadas, dependiendo de la clase del anticuerpo. Mediante la unión a proteínas específicas la región Fc se asegura que cada anticuerpo genera una respuesta inmune apropiada para un antígeno dado. La región Fc también se une a varios receptores celulares como el receptor del Fc y otras moléculas del sistema inmunitario como las proteínas del complemento. Al efectuar esto, media en diferentes efectos fisiológicos incluyendo la opsonización, lisis celular y desgranulación de las células cebadas, basófilos y eosinófilos
Los anticuerpos pueden presentarse en distintas variedades conocidas
87945 Roy Rodriguez Panesso
ResponderEliminarAntigenicidad
Capacidad para dar lugar a la producción de anticuerpos. El grado de antigenicidad depende del tipo y cuantía de la sustancia concreta, de las circunstancias del huésped y de su grado de sensibilidad al antígeno y de su capacidad para producir anticuerpos
antigenia, o antigénico (potencial). Poder de comportarse como un antígeno, es decir, de provocar la formación de anticuerpos. Para ciertos autores, la antigenicidad sería la facultad, para el antígeno, de modificar el comportamiento inmunológico del organismo en el cual se ha introducido, sin que por ello haya forzosamente producción de anticuerpos, por ejemplo, provocando un estado de tolerancia inmunitaria. Difiere así del poder inmunógeno.
87945
87945 Roy Rodriguez Panesso
ResponderEliminarEpitopos
Es la unidad más pequeña de un antígeno que puede unirse a un BCR o TCR específico, o a los anticuerpos
secretados. Es decir, los epitopos son las regiones inmunológicamente activas
de un antígeno. Pueden ser lineales (continuos) o conformacionales.
El sistema inmunológico humano es capaz de reconocer un amplio rango de distintos epítopos. Los anticuerpos son proteínas que produce el sistema inmune que reaccionan con su antígeno meta (que generalmente son proteínas presentes en organismos que invaden o materiales extraños tales como proteínas alimentarias). Si pensamos que los anticuerpos son llaves que se corresponden a una cerradura meta ubicada en el antígeno, el epítopo sería la ranura que corresponde a una llave específica. El epítopo suele ser un parche de residuos de aminoácidos, los bloques que forman las proteínas. El epítopo es la ubicación en el antígeno en donde la molécula del antígeno atraca, como una llave que abre su cerradura.
87945
87945 Roy Rodriguez Panesso
ResponderEliminarEstructuras de los anticuerpos
La estructura de los anticuerpos está basado en la cadena peptídica, una secuencia de aminoácidos sucedidos uno tras otro y unidos por sus funciones amino y carboxilo. Esta gran cadena posee distintos niveles de organización: primario, secundario, terciario e incluso cuaternario. están formados por dos subunidades, las cuales a su vez están compuestas por otras dos subunidades, una cadena liviana y una cadena pesada. Ambas cadenas se subdividen en una región constante y en una región variable. Esta última es la responsable de la unión del anticuerpo con el antígeno mientras que la región constante puede tener distintas funciones dependiendo del dominio. Existen distintos tipos de cadenas pesadas (mu, epsilon, alfa, gamma, delta) y distintos tipos de cadenas livianas (kappa y lambda), los cuales se encuentran determinados por la secuencia de aminoácidos de las regiones constantes.Se ha podido determinar dos zonas dentro de las inmunoglobulinas, ambas generadas a base de experimentos con enzimas. Un primer fragmento es el fragmento Fab (que puede unirse a los antígenos) y un segundo fragmento es el Fc (que no puede unirse a los antígenos pero sí puede servir como ligando de receptores Fc).Además, dentro de la región variable existen zonas más variables y regiones más conservadoras. Las primeras se denominan CDR y son 3, CDR1 , CDR 2, CDR 3. Esta última es la más variable.
87945
87945 Roy Rodriguez Panesso
ResponderEliminarContinuacion de estructura de las inmunoglobulinas
Cadenas Ligeras.
Hay dos tipos de cadenas ligeras, estructuralmente diferentes, que se conocen como cadenas ligeras tipo kappa (k) y cadenas ligeras tipo lambda (l). La familia de genes que codifica para la cadena ligera k se localiza en el cromosoma 2 y los loci de los genes homólogos que codifican para la cadena l, en el cromosoma 22. En cada molécula de inmunoglobulina las dos cadenas ligeras son del mismo tipo, k o bien l, pero nunca existe una de cada tipo en la misma inmunoglobulina.
Las cadenas ligeras están formadas por unos 200 aminoácidos con la particularidad de que existen dos puentes disulfuro que unen grupos de unos cincuenta aminoácidos. Concretamente la IgG1 posee 214 aminoácidos y su estructura secundaria y terciaria están determinadas por dos puentes disulfuro intracatenarios que unen los aminoácidos 23 con el 88 y 134 con el 193 . A su vez, estas cadenas ligeras tienen otro puente disulfuro intercatenario, por el cual cada una de ellas se une a una cadena pesada para constituir la unidad básica de las inmunoglobulinas. Este puente se encuentra en el último aminoácido (214) de la parte carboxílica para el tipo k y en el penúltimo para el tipo l.
Cadenas pesadas.
Estas cadenas poseen unos cuatrocientos aminoácidos estableciéndose entre algunos de ellos puentes disulfuro (intracatenarios) que asocian unos 60 aminoácidos y que condicionan la estructura secundaria del polipéptido. Por ejemplo, las cadenas pesadas de la IgG1 poseen 440 aminoácidos y los puentes disulfuro unen el aminoácido 22 con el 96, el 144 con el 200, el 261 con el 321 y el 367 con el 425.
Estas dos cadenas pesadas están unidas la una a la otra por puentes disulfuro intercatenarios, ya indicados anteriormente, y que pueden ser de uno a cinco dependiendo del tipo de inmunoglobulina.
En estas cadenas pesadas, y a nivel de los puentes disulfuro intercatenarios, hay una zona de unos 15 aminoácidos, de gran flexibilidad debido a su estructura y constituye lo que se denomina zona bisagra por donde se deforma la molécula de inmunoglobulina cuando se produce la unión con el antígeno, facilitándose así su acoplamiento con éste. Los loci de los genes que codifican para la cadena pesada se localizan en el brazo largo del cromosoma 14.
87945
87945 Roy Rodz Panesso
ResponderEliminarIsotipo de inmunoglobulinas
La información genética que permite la existencia de un repertorio amplio de anticuerpos reside en diferentes cromosomas: los genes que codifican para las diferentes regiones e isotipos de cadenas pesadas se encuentran en el cromosoma 14, los que codifican para la cadena ligera kappa se encuentran en el cromosoma 2 y en el 22, los que codifican para la cadena lambda. La distribución de los antisuerpos varía entre la vida prenatal y postnatal temprana (predominio de anticuerpos IgG maternos) y el resto de la vida del ser humano (predominio de los anticuerpos IgM en la respuesta inmune primaria y predominio global de los IgG en suero y de los IgA en secreciones mucosas, dentro de la respuesta secundaria). IgG: Es la inmunoglobulina de mayor concentración en circulación sanguínea (70% del total de los anticuerpos). Es un monómero, donde la cadena pesada puede ser de 4 subclases distintas. Es el anticuerpo preponderante en la respuesta secundaria. Todas las subclases atraviesan la placenta y todas, excepto la IgG4 pueden activar el complemento. Su localización puede ser tanto intravascular como extravascular. IgM: Es un anticuerpo que circula en sangre como un pentámero de la unidad básica. La molécula posee además una cadena adicional, llamada J, que parece desempeñar un papel importante en la polimerización de las subunidades. La IgM es el anticuerpo predominante en las respuestas primarias. La IgM encontrada en la superficie de los linfocitos B (BCR) es monomérica. IgA: Supone el 15-20% del total de imunoglobulinas, pero la mayoría (> 80%) circulan en el ser humano como un monómero de la unidad básica. Sin embargo, se encuentra en gran cantidad en las secreciones del tracto respiratorio, genitourinario, saliva, lágrimas, calostro y leche materna. En estas secreciones adopta una estructura dimérica que, al igual que la IgM, posee también una cadena J. Para poder atravesar los epitelios se une a un receptor de las células epiteliales que en un momento dado (después del transporte a la porción apical de la célula) sufre un clivaje y queda enganchado al dímero. A este fragmento de receptor, o cadena adicionada, se le denomina componente o pieza secretora y confiere al anticuerpo mayor resistencia al ataque proteolítico. Existen dos subclases de IgA, llamadas IgA1 e IgA2 funcionalmente equivalentes. IgD: Posee estructura monomérica. Constituye menos del 1% del total de anticuerpos circulantes, pero está presente, al igual que la IgM monomérica, en la superficie de todos los linfocitos B maduros vírgenes (BCR). Aunque se desconoce su función precisa, se sospecha que participa en la activación y/o diferenciación del linfocito B después del reconocimiento antigénico, a partir del cual cesa su expresión en la membrana. IgE: Posee una estructura monomérica. Es el anticuerpo de más baja concentración en circulación, pero se halla fijado (a través de su porción Fc) en la superficie de células cebadas y basófilos tisulares. Es el responsable de los fenómenos de hipersensibilidad inmediata y parece desempeñar un papel importante en la respuesta inmune contra ciertos parásitos (helmintos).
Los anticuerpos reconocen al antígeno en su forma nativa y su unión se produce a través de los epitopos o determinantes antigénicos, por un lado, y las porciones hipervariables de las regiones variables de los anticuerpos. Las fuerzas que mantienen la unión no son covalentes (puentes de hidrógeno, interacciones electrostáticas, interacciones hidrófobas y fuerzas de Van der Waals) y esta unión mantiene diferentes grados de afinidad y avidez.
87945 Roy Rodriguez Panesso
ResponderEliminarIgA
es la clase predominante de anticuerpo en las secreciones seromucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica.1 (IgA secretora)
Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas.
La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 ml
87945
87945 Roy Rodz Panesso
ResponderEliminarIgG
es una de las cinco clases de anticuerpos humorales producidos por el organismo. Se trata de la inmunoglobulina predominante en los fluidos internos del cuerpo, como son la sangre, el líquido cefalorraquídeo y el liquido peritoneal (líquido presente en la cavidad abdominal). Esta proteína especializada es sintetizada por el organismo en respuesta a la invasión de bacterias, hongos y virus. Es la inmunoglobulina más abundante del suero, con una concentración de 600-1.800 mg por 100 mL.1 La IgG constituye el 80% de las inmunoglobulinas totales.
La IgG es la única clase de inmunoglobulinas que atraviesa la placenta, transmitiendo la inmunidad de la madre al feto de manera natural y pasiva. Es la inmunoglobulina más pequeña, con un peso molecular de 150.000 Daltons,1 así puede pasar fácilmente del sistema circulatorio del cuerpo a los tejidos. La síntesis de IgG se controla principalmente por el estímulo de los antígenos. En el caso de animales axénicos (sin microbios), con niveles de IgG muy bajos, el nivel de IgG se eleva en cuanto se les traslada a un ambiente normal.
Hay veces que estas inmunoglobulinas no responden adecuadamente. Especialmente en casos de Anaplasmosis simple.
Las cadenas H de la IgG son del isotipo γ, contiene un 2-3% en peso de glúcidos.
87945
87945 Roy Rodz Panesso
ResponderEliminarIgM
constituyendo un 6% de la población presente en sangre. También se encuentra presente en elasmobranquios, teleósteos, anfibios, reptiles y aves, siendo uno de los anticuerpos más antiguos en la historia evolutiva1 . Se denomina también macroglobulina (de ahí el nombre de la enfermedad en la que se presenta exceso, macroglobulinemia de Waldenström) debido a su tamaño: es la inmunoglobulina más grande (950.000 Daltons),2 aunque el tamaño no se debe exclusivamente al peso molecular real de la molécula, sino que ésta presenta la capacidad, a través de su región Fc, de interaccionar con otras cuatro moléculas de IgM, formando un complejo de alto peso molecular de cinco moléculas de IgM.
La capacidad de la IgM para formar estos complejos -lo cual le da gran facilidad para unir el complemento- es la que le da el poder de opsonizar determinados antígenos, provocando la lisis de bacterias, envueltas víricas y otros agentes patógenos. Es el primer tipo de inmunoglobulina sintetizada en respuesta a una infección.
87945
87945 Roy Rodz Panesso
ResponderEliminarIgA
es la clase predominante de anticuerpo en las secreciones seromucosas del organismo como saliva, lágrimas, calostro, leche y secreciones respiratorias, gastrointestinales y genitourinarias. En sangre, se encuentra como una molécula monomérica, pero en las mucosas se encuentra en forma dimérica.1 (IgA secretora)
Actúan como la defensa inicial contra los patógenos invasores (virus y bacterias) antes de que penetren en el plasma; identifican los antígenos patógenos e impiden que se instalen en las mucosas.
La IgA tiene una masa molecular que oscila entre 170.000 y 720.000, ya que forma estructuras poliméricas de la unidad estructural básica; la cadena H es del isotipo α, contiene un 7-12% en peso de glúcidos y su concentración en el suero es de 90-420 mg por 100 mL.
87945
87945 Roy Rodz Panesso
ResponderEliminarIgD
Se halla en cantidades pequeñas, 0-1% de las inmunoglobulinas, y tiene un peso de 185.000 Daltons. No es secretada por los plasmocitos. Se conoce por ser el mayor componente de la superficie de muchos linfocitos B en etapas de maduración. Su presencia sobre las células B sirve como marcador de diferenciación, y puede servir para controlar la activación y supresión de linfocitos. La IgD no se encuentra de manera soluble en el plasma.
Está presente en la superficie de la mayor parte de linfocitos B circulantes, indicando que las células B vírgenes están listas para entrar en contacto con el antígeno. La IgD se pierde durante la estimulación antigénica, las células de memoria han perdido esta inmunoglobulina.
La IgD es una proteína funcional importante, pero su papel no es bien conocido; se sugiere que es un receptor antigénico de membrana, que conduciría a la diferenciación linfocitaria y también que es un ligando para los receptores de IgD en la inmunoregulación de las células T ‘helper’. Es muy susceptible a la proteólisis.
La IgD representa menos del 1% del total de inmunoglobulinas plasmáticas; la concentración en suero depende de la edad y de la herencia genética. En suero de pacientes con mieloma IgD se encuentran concentraciones de IgD muy elevadas.
87945
87945 Roy Rodz Panesso
ResponderEliminarGenetica de las inmunoglobulinas
El sistema inmune es capaz de producir hasta 108 moléculas de Ac cada una con especificidad única para Ag. El mecanismo genético único para generar diversidad de Ac fue descubierto al estudiar genes codificantes de las proteínas de mieloma con la línea ADN germinal. Cada linfo B rearregla su ADN para un gen funcional de cadena H y uno de cadena L.
La especificidad de Ac se determina por la secuencia de aa de los dominios V de sus cadenas H y L, donde se capta Ag.
La secuencia de la región C es la misma para las cadenas H y L de un isotipo determinado y no tiene efecto en la especificidad.
La familia de Ig tiene un número pequeño de dominios de C, unidos en varias combinaciones con una variedad casi ilimitada de secuencias de la región V.
La información está dispersa a lo largo del ADN, en múltiples segmentos codificadores (exones), separados por regiones ADN no codificador (intrones), cuando el gene se transcribe al ARN, los intrones se eliminan y los exones se unen.
87945 Roy Rodz Panesso
ResponderEliminarContinuacion de Genetica de las inmunoglobulinas
Genes para cadenas ligeras
La información genética necesaria para producir cadenas k se encuentra en un solo locus (k) en el cromosoma 2.
Las cadenas l, nacen de un complejo genético similar en el cromosoma 22; la unión de segmentos Vl y Jl se presentan de una manera idéntica a los segmento k.
Las cadenas H se derivan de una sola región del cromosoma 14; cada región C de la cadena H está codificada por un grupo de varios exones cortos.
La secuencia de la región C (CH) para cada uno de los nueve isotipos de la cadena pesada, se rearreglan a lo largo del cromosoma; sólo esta presente una copia de cada uno.
87945
87945 Roy Rodz Panesso
ResponderEliminarCalostro
es un líquido secretado por las glándulas mamarias durante el embarazo y los primeros días después del parto, compuesto por inmunoglobulinas, agua, proteínas, grasas y carbohidratos en un líquido seroso y amarillo.
"Para el nacimiento del bebé se preparara uno de los alimentos más importantes de su vida, el calostro. Este alimento es la primera leche que se produce y su duración está entre los 2 y 5 días, antes de que se empiece a producir la leche definitiva.
El calostro es un alimento ideal y primordial, ya que en él está la primera inmunización del bebé y resuelve además las necesidades alimentarias de sus pequeños órganos, aún no maduros.
Algunas diferencias existentes con respecto a la leche definitiva son por ejemplo la densidad, mucho mayor y más pegajoso. Por su color, al calostro lo apodan oro líquido, esto se debe a su color algo dorado, claro que depende de la mama que lo produce, a veces es más oscuro y otras veces más claro.
Precalostro. Antes de hablar del calostro previamente hay que saber que existe el precalostro que se forma durante el tercer trimestre de gestación, los pechos empiezan a secretar un líquido que se encuentra formado por plasma, sodio, cloro, inmunoglobulinas, lactoferrina, seroalbúmina y una ínfima cantidad de lactosa. Este líquido se llama Precalostro y avisa que los pechos están preparándose para amamantar al nuevo bebé.
87945